Antropología
Teorías de la Evolución Humana
EVOLUCIÓN BIOLÓGICA
I. INTRODUCCIÓN.
El mundo y la sociedad en que vivimos, cambia constantemente, todo lo que nos rodea es frágil y susceptible a una cambio o transformación inmediata. Las más desarrolladas teorías son desbaratadas gracias a los altos avances científicos, la más grande inversión en computación puede desevaluarse en una semana, cada día podemos ver más lejos estudiando nuestro universo y cada día más internamente en nuestras microestructuras. Al parecer “cambia todo cambia” y “Evoluciona todo evoluciona”, ¿pero entonces es que nos hemos quedado atrás? todo a nuestro alrededor cambiando, “mutando”, evolucionando, ¿pero nosotros?, ¿evolucionamos?, ¿cambian nuestras formas biológicas y las de las demás especies? estas preguntas se plantean en el mismo campo del desconocido mundo del origen de la vida. Algo tan lento donde perdemos parte del proceso más preciado de la ciencia: la observación. El origen y la diversificación de la vida ocurrieron hace millones de años, son procesos largos y difíciles de comprender, muchas veces se mueven más en el campo de la especulación. Y es aquí donde gran cantidad de brillantes científicos han unido fuerzas para contestar a las interrogantes de la evolución y la vida. Y es el objetivo de este documento dar a conocer los estudios que se han hecho a lo largo de nuestra historia desde el análisis de pruebas de hace millones de años atrás hasta las teorías más recientes como la neutralista; las teorías que responden a nuestras interrogantes, y las pruebas de las cuales se aferran estos hombres detrás de estas teorías para comprobar sus “hazañas”. A continuación las teorías por las cuales el hombre puede jactarse de ser “más evolucionado”.
II. DEFINICIÓN DE CONCEPTO.
“La evolución es un cambio genético en una población, vale decir, en organismos de una misma especie, que no se refiere a los cambios sufridos por un individuo durante su vida. Los individuos no evolucionan, pero sí lo pueden hacer las poblaciones. Mediante el concepto de evolución, los biólogos están capacitados para encontrarle un sentido a la impresionante cantidad en variedad de seres vivos que conviven en el planeta”
III. TEORÍAS.
“En esta tercera parte incluimos a siete autores de teorías de la evolución desde
la teoría de la panspermia hasta la neutralista, y explicamos en qué consiste su
planteamiento, abarcando casi un siglo de descubrimientos”
1. Svante Arrhenius:
Después de quedar refutada la teoría de origen espontáneo, que explicaba que las especies habían surgido de manera espontánea a partir de la materia inanimada, surgió una nueva pregunta ¿Si los seres vivos no pueden desarrollarse a partir de materia inanimada, entonces cómo comenzó la vida en nuestro planeta, millones de años atrás?
Uno de los primeros en intentar responder a esta interrogante fue el químico sueco Svante A. Arrhenius (1859-1927), que propuso que los gérmenes de vida (esporas) habrían llegado del espacio exterior en meteoritos que se habrían desprendido de un planeta en el que ya habría vida traídos a la tierra por la presión de la luz.
Ideas de este tipo han surgido una y otra vez a lo largo de la historia. Sin embargo, dejan sin resolver el problema del origen de la vida, dado que los procesos que explican su establecimiento, ya sea en la Tierra o en el espacio, no son abordados sino desplazados a un tiempo y lugar inaccesibles. Quizás las condiciones interiores del meteorito, hayan sido propicias para el desplazamiento, pero deja abiertas cuestionantes como qué habría pasado en la explosión contra la tierra. Además actualmente esto serviría para explicar el origen de la vida, pero no cómo evolucionan.
2. George Cuvier:
La fuerza dominante en la ciencia europea a principios del siglo XIX era George Cuvier (1769-1832). Cuvier fue el fundador de la paleontología de los vertebrados, el estudio científico del registro fósil de los animales con esqueleto interno, y de la anatomía comparada. Experto en anatomía y zoología, aplicó su conocimiento del modo en que los animales estaban construidos al estudio de los animales fósiles y era capaz de hacer deducciones brillantes acerca de la forma de un animal completo a partir de unos pocos fragmentos óseos.
Actualmente consideramos que la paleontología y la evolución están tan íntimamente conectadas que nos sorprende saber que Cuvier fue un influyente y firme adversario de las teorías evolutivas. A pesar de sus profundos conocimientos acerca de los seres vivos actuales y extintos, Cuvier consideraba que las especies habían sido creadas simultáneamente por un acto sobrenatural o divino y que, una vez creadas, se mantuvieron fijas o inmutables. Esta postura que se conoce como "fijismo" era predominante en el pensamiento de los naturalistas de la época.
Estableció a ley de correlación de las partes y una clasificación animal con cuatro tipos independientes: vertebrata, articulata (artrópodos y gusanos segmentados), mollusca (moluscos e invertebrados con simetría bilateral), radiata (cnidarios y equinodermos). Para Cuvier, estas ramas eran completamente diferentes entre ellas y no podían estar conectadas por ninguna transformación evolutiva. Las similitudes orgánicas eran debidas al desempeño de funciones comunes y no a la existencia de un antepasado común. Por lo tanto, para Cuvier, la función determina la forma.
3. Jean-Baptiste de Lamarck:
El primer científico moderno que elaboró un concepto sistemático de la evolución fue Jean Baptiste Lamarck (1744-1829), si bien no fue el primero en poner en duda la idea de que las especies son fijas y no cambian. Este "naturalista justamente célebre", como el propio Darwin (1809-1882) lo calificó, propuso audazmente en 1801 que todas las especies, incluyendo al Homo sapiens, descienden de otras especies. Fue su largo estudio de los organismos unicelulares y los invertebrados lo que lo llevó a considerar a los seres vivos desde el punto de vista de una complejidad en continuo aumento, y a cada especie como derivada de una más primitiva y menos compleja.
A diferencia de Cuvier, que adhería a las ideas fijistas, Lamarck interpretó estas evidencias como si las formas más complejas hubiesen surgido de las formas más simples por una suerte de progresión. De acuerdo con su hipótesis, esta progresión depende de dos fuerzas principales. La primera es la "herencia de los caracteres adquiridos". Los órganos en los animales se hacen más fuertes o más débiles, más o menos importantes, por su uso o su desuso y por la influencia del medio ambiente, y estos cambios, se transmiten a la descendencia.
La segunda fuerza igualmente importante en el concepto de evolución de Lamarck fue un principio creador universal, un esfuerzo inconsciente y ascendente en la Scala Naturae, que impulsaba a cada criatura viva hacia un grado de complejidad mayor. Algunos organismos podían quedar apartados; el orangután, por ejemplo, había sido desviado de su curso al ser atrapado por un ambiente desfavorable, pero la "voluntad" estaba siempre presente.
El hecho de que Lamarck no pudiera comprobar la teoría de los caracteres adquiridos, ya que para esto se necesitan recursos tecnológicos de los que carecía, junto con que la “voluntad” sea un hecho metafísico, hizo que su teoría careciera de credibilidad y que Cuvier atacara sus ideas constantemente; pero el aporte que este científico hace a la teoría evolucionista, es importantísimo, ya que es el primero en postular el progresismo (contrapuesto al fijismo), introducir el factor de la herencia y lo que veremos más adelante como órganos rudimentarios.
4.Charles Darwin y Alfred Russel Wallace:
En su viaje que incluyó las islas Galápagos, Darwin vio a varias especies de pinzones muy parecidas, cosa que le hizo sospechar que estas especies provenían de la misma. Otro factor que impactó a Darwin e infundió aun más sospechas fueron los fósiles encontrados en América del Sur. En su gran mayoría, los fósiles pertenecían a la misma familia de las especies vivas. Todo esto le hizo plantearse serias dudas acerca de la inmutabilidad de las especies. Podemos decir que el origen de su teoría proviene de dos puntos claves; su viaje en el Beagle y el conocimiento de la teoría de Malthus sobre el crecimiento de la población en relación a los recursos.
En 1858, A. R. Wallace le envió un escrito en el que proponía una teoría similar a la de Darwin. Wallace había llegado a las mismas conclusiones que él. Publicaron un juntos, en el que exponían la teoría evolucionista de la selección natural llamado “El origen de las Especies” publicado en 1859.
Ya en su viaje alrededor del mundo, Darwin había detectado la evolución orgánica, pero no sabía como funcionaba. La respuesta la encuentra en las variaciones entre individuos de una misma especie. El criador de alguna especie doméstica es capaz de escoger a los más capacitados para que procreen y así conseguir perfeccionar una especie para su mejor uso (por ejemplo los gallos de pelea). Darwin observa que si el hombre es capaz de hacer variar a una especie, aunque sólo sea exteriormente, la naturaleza debe ser mucho más capaz. Darwin deriva de la selección artificial hecha por el hombre la selección natural.
Pero para poder elegir entre varios animales es necesario que haya muchos ejemplares. Es aquí donde Darwin acude a la teoría de Malthus. La teoría malthusiana de que la población crece ascendentemente, mientras que los recurso decrecentemente es aplicada al resto de los seres vivos. Así, la sobreproducción de seres vivos haría que los menos aptos perecieran y que sólo sobrevivieran los mejor dotados, los más aptos, los mejor adaptados a su entorno. Así pues, las variaciones dentro de una misma especie supone el ser apto o no para su supervivencia. Si estas variaciones fuesen tan sólo personales y no heredables, la teoría de Darwin no tendría ningún sentido. Es por ello que Darwin habla de que los más aptos “tendrán seguramente las mejores probabilidades de conservarse en la lucha por la vida, y, por el poderoso principio de la herencia, éstos tenderán a producir descendencia con caracteres semejantes.”
El concepto original de Darwin y de Wallace acerca de cómo ocurre la evolución todavía sigue proporcionando el marco básico para nuestra comprensión del proceso. Ese concepto se funda en cinco premisas:
Los organismos engendran organismos similares; en otras palabras, hay estabilidad en el proceso de la reproducción.
En la mayoría de las especies , el número de individuos que sobreviven y se reproducen en cada generación es pequeño en comparación con el número total producido inicialmente.
En cualquier población dada ocurren variaciones aleatorias entre los organismos individuales, algunas de las cuales son hereditarias, es decir, que no son producidas por el ambiente.
La interacción entre estas variaciones hereditarias, surgidas al azar, y las características del ambiente determinan en grado significativo cuáles son los individuos que sobrevivirán y se reproducirán y cuáles no. Algunas variaciones permiten que los individuos produzcan más descendencia que otros. Darwin llamó a estas características variaciones "favorables" y propuso que las variaciones favorables heredadas tienden a hacerse cada vez más comunes de una generación a otra. Este es el proceso al que Darwin llamó selección natural .
Dado un tiempo suficiente, la selección natural lleva a la acumulación de cambios que provocan diferencias entre grupos de organismos.
Por lo tanto, la selección natural marca la pauta de la evolución de las especies. Las variaciones dadas en los individuos de cada especie y que marcaran su aptitud o no, vienen dadas por diversos factores. Según Darwin “la ignorancia de las leyes de variación es profunda”. Aún así, se aventura a explicar las causas de las variaciones de las especies. Por un lado argumenta que el cambio de condiciones produce normalmente variaciones fluctuantes. Por otro lado cree que una causa poderosa para la variación es el uso y el desuso de determinados miembros del organismo, que con el paso del tiempo supondrá el desarrollo de la parte usada y la inutilidad o desaparición de la parte no usada (por ejemplo las alas y las patas de las avestruces). La especie que haya sufrido la variación y que procree muchos descendientes que conserven la modificación habrá logrado mantener fijo el órgano variado.
Para corroborar que está en lo cierto, Darwin cree que se puede hablar de diferentes especies y de variedades dentro de las especies. Las variedades de una misma especie son fecundas entre sí, y pueden dar lugar a una variedad totalmente distinta de la de sus progenitores, pero que forma parte de la misma especie. Sin embargo, si se trata de unir a dos especies diferentes, la diferencia sexual no deja sino lugar al nacimiento de lo que Darwin llamó híbrido. Un híbrido acostumbra a ser estéril, aunque no se trata de una ley universal, pues Darwin ha comprobado que hay animales domésticos de diferentes razas que al cruzarse son totalmente fecundas. El problema con el que se encuentra Darwin en este punto es que desconoce el por qué surgen algunos híbridos estériles y otros fecundos. Su teoría flojea fuertemente en este punto. Darwin aún no podía conocer el funcionamiento de la genética y por lo tanto de la herencia genética traspasada de padres a hijos. Las variaciones se producirían al azar, en tanto que su causa reside en un proceso natural independiente de las exigencias ambientales que se le planteaban a la especie.
Aun así, Darwin muestra que la idea del creacionismo es una idea errónea, pues no se trata de la creación de especies diferentes desde un principio, sino de variedades clasificadas en especies, subgéneros y finalmente géneros. Si anuviéramos atrás en el tiempo, según Darwin, veríamos que todas las especies de las que hoy tenemos conocimiento, son especies que provienen de un grupo más reducido: “Cuando considero a todos los seres no como creaciones especiales, sino como los descendientes directos de unos cuantos seres que vivieron mucho antes de que se depositase la primera capa del sistema cámbrico, me parece que se ennoblecen mis ojos”.
Darwin acaba su teoría especulando de un futuro prometedor, pues la selección natural sólo elige a los más aptos y por tanto “ todos los dones corporales e intelectuales tenderán a progresar hacia la perfección.” El mundo futuro estará formado por seres más perfectos, más evolucionados, y esta idea fascinó a Darwin hasta sus últimos días.
5. Hugo de Vries:
En 1903, el botánico holandés Hugo de Vries (1848-1935) observó que en las poblaciones naturales aparecían ocasionalmente individuos que diferían en alguna característica del resto de los ejemplares de la población y que eran capaces de producir descendientes iguales a sí mismos. Sobre la base de estos hallazgos propuso que los genes sufrían alteraciones súbitas e independientes del medio ambiente, a las que llamó mutaciones, que podían ser transmitidas a las siguientes generaciones. Así, la mutación pasaba a ser el mecanismo por el cual surgen nuevas variantes en los genes, a partir de errores al azar que ocurren en el material genético. Con esta nueva postura a lo que Darwin llamaba variación, quedaba finalmente superada la concepción de la herencia de los caracteres adquiridos.
Sin embargo, estas conclusiones no se integraron al marco teórico del darwinismo. Los genetistas propusieron que las mutaciones producirían efectos tan drásticos en los organismos, que llegarían incluso a originar nuevas especies. En otras palabras, las mutaciones no fueron consideradas como materia prima de la selección natural, sino como un mecanismo alternativo capaz de explicar por sí mismo la evolución de las especies. Así, por unos 30 años, la teoría mutacionista representó una hipótesis alternativa a la selección natural para explicar la evolución biológica.
6. Teoría Sintética de la Evolución:
Una de las principales debilidades de la teoría de la evolución, según fuera formulada por Darwin, era la ausencia de un mecanismo válido para explicar la herencia . El desarrollo posterior de la genética permitió dar respuesta a tres cuestiones que Darwin nunca pudo resolver:
1) ¿de qué manera se transmiten las características heredadas de una generación a la siguiente?;
2) ¿por qué las características heredadas no se "mezclan", sino que pueden desaparecer y luego reaparecer en generaciones posteriores y
3) ¿de qué manera se originan las variaciones sobre las cuales actúa la selección natural ?
La combinación de la teoría de la evolución de Charles Darwin (1809-1882) con los principios de la genética mendeliana se conoce como la síntesis neodarwiniana o la Teoría Sintética de la evolución. Esta teoría intenta relacionar la Teoría de la evolución con la paleontología, la sistemática y la genética. Los principales representantes de las síntesis fueron el genetista Theodosius Dobzhansky (1900-1975), el zoólogo Ernst Mayr , el paleontólogo George G. Simpson (1902-1984) y el botánico George Ledyard Stebbins (1906-), todos ellos de Estados Unidos, y el zoólogo Julian Huxley (1887-1975) de Inglaterra.
Dobzhansky propuso que la evolución puede percibirse como un cambio de frecuencias génicas en el seno de una población. Mayr propuso, en su libro Sistemática y origen de las especies, dos nociones que permiten comprender cómo se forman las nuevas especies: el concepto biológico de especie y el modelo de especiación geográfica. Simpson aplicó a los fósiles las ideas de Dobzhansky sobre la evolución. Para los defensores de la Teoría Sintética, la evolución de las especies resulta de la interacción entre la variación genética que se origina en la recombinación de alelos y las mutaciones, y la selección natural.
Aunque en la actualidad, los esfuerzos en la investigación acerca de la evolución se centran en descubrir cuáles son y como funcionan los mecanismos evolutivos, la síntesis darwiniana describe tres mecanismo graduales de acción. La microevolución y la macroevolución, ambas están divididas en el concepto de especie.
Gracias a la teoría sintética se explica la microevolución, que son pequeños cambios que se van acumulando en los individuos de una población por mecanismos de recombinación genética de alelos (genes) o por cruzamiento de variedades (poblaciones dentro de una especie que pueden reproducirse) dentro de una especie, se dan mutaciones genéticas, las que aún no se sabe exactamente por qué se originan. Esta mutaciones producen cambios en esa especie. Las evoluciones que abarca la microevolución se realizan siempre por debajo el concepto de especiación.
Las mutaciones pueden ser favorables o desfavorables, en el caso que sean favorables, por selección natural, será la mutación más apta la que sobreviva y pueda transmitirse a las siguientes generaciones. Este proceso continua, siguen produciéndose cambios en las variedades, entonces se llaman razas. En estas razas sigue habiendo cambios, entonces se llaman subespecies. Siguen produciéndose cambios, entonces puede ocurrir que se produzca una barrera geográfica, reproductiva, etc y aísle a las poblaciones durante un tiempo, entonces, evolucionan por caminos diferentes.
En muchos casos, los grupos de organismos que se separan de la población original, y quedan aislados del resto, pueden alcanzar una diferenciación suficiente como para convertirse en una nueva especie. Este proceso, denominado especiación , ha ido ocurriendo durante 3.800 millones de años, dando origen a la diversidad de organismos que han poblado la Tierra en el pasado y en la actualidad. Una especie , desde un punto de vista biológico, es un grupo de poblaciones naturales cuyos miembros pueden cruzarse entre sí, pero no pueden hacerlo -o al menos no lo hacen habitualmente con los miembros de poblaciones pertenecientes a otras especies. En este concepto, el aislamiento en la reproducción respecto de otras especies es central. ver cuadro (1)
La macroevolución se ocupa del estudio del proceso evolutivo a gran escala es decir, el análisis de los patrones generales del cambio evolutivo a través del tiempo geológico, y se centra en el estudio de los procesos evolutivos que ocurren por encima del nivel de especie , e intenta explicar el por qué de los grupos taxonómicos mayores.
El análisis del registro fósil revela diversos patrones de cambio macroevolutivo: la evolución convergente, la evolución divergente, el cambio filético , la cladogénesis , la radiación adaptativa y la extinción. La evolución convergente produce adaptaciones notablemente similares en organismos lejanamente relacionados, mientras la evolución divergente conduce al establecimiento de adaptaciones ampliamente diferentes en organismos íntimamente relacionados. El cambio filético es el cambio gradual dentro de un linaje individual a lo largo del tiempo. Por el contrario, la cladogénesis es el cambio evolutivo que produce la bifurcación de poblaciones unas respecto de otras para formar especies nuevas, originando nuevas ramas. La radiación adaptativa es la formación rápida de muchas especies nuevas a partir de un único ancestro, las cuales son capaces de invadir nuevas zonas adaptativas por poseer una nueva característica clave.
La extinción es la desaparición completa de una especie. El registro fósil muestra una tasa de extinción de fondo lenta y continua, interrumpida periódicamente por extinciones masivas que involucran a enormes números de especies y, en ocasiones, grupos de rango superior. La macroevolución puede ser interpretada como el resultado de una combinación de estos patrones.
Los paleontólogos S. Gould y N. Eldredge han presentado evidencias de un patrón adicional de macroevolución, conocido como modelo de los equilibrios intermitentes. Este modelo propone que las nuevas especies se forman en períodos muy breves, casi instantáneos en la escala del tiempo geológico, a partir de pequeñas poblaciones periféricas aisladas. Estas nuevas especies, en ocasiones, compiten con éxito con las especies previamente existentes, que pueden llegar a extinguirse. La ausencia de cambio gradual en el registro fósil evidencia que una vez establecidas, las especies persisten durante períodos prolongados con poco cambio (estasis), hasta que se extinguen abruptamente. Los defensores del equilibrio intermitente proponen que los cambios principales en la evolución ocurren como resultado de la selección que actúa sobre las especies en el nivel macroevolutivo, así como la selección natural lo hace sobre los individuos, en el nivel microevolutivo.
7. Teoría Neutralista:
A escala molecular existe un apoyo creciente a la idea de que la mayoría de las variaciones evolutivas son en realidad neutrales. Esta teoría neutral de la evolución ha sido defendida por el distinguido genetista japonés Motoo Kimura. La teoría neutral no afirma que los genes no estén realizando algo útil, más bien sugiere que formas diferentes del mismo gen son indistinguibles en cuanto a sus efectos. Por ello, una mutación de una forma de un gen a otra es neutral en cuanto a que la modificación no afecta al fenotipo. Kimura y sus colaboradores apuntaron la evidencia de que la mayoría de las sustituciones de los genes en la naturaleza era neutral. Es decir, bajo su punto de vista, es la principal causa de variación genética en las poblaciones.
La teoría neutral se describe algunas veces como antidarwiniana, aunque ello es un gran error. Las mutaciones neutrales son equivalentes a los cambios experimentados por la tipografía desde la época de los romanos hasta el derecho romano actual: el significado de las frases escritas es invariable. La selección darwiniana juzga a los genes por su expresión fenotípica (por el significado de sus frases). Si una mutación carece de efectos sobre el fenotipo (es puramente un cambio de tipografía), la selección natural será indiferente a ésta. La teoría neutral no se pronuncia, de una forma o de otra, acerca de la importancia de la selección darwiniana a nivel de los fenotipos (fenotipo: Características observables de un organismo que resultan de las interacciones entre su concepción original y el ambiente).
Una consecuencia interesante de ésta teoría es la idea de un “reloj genético molecular”. Si la mayoría de las sustituciones genéticas son neutras, es de esperar que la tasa de sustituciones sea más o menos constante para cualquier gen determinado. Suponiendo esto, la época en la que vivió el antecesor común de cualquier pareja de especies se puede calcular a partir del número de diferencias en los aminoácidos entre ambas especies. Al principio, dichas fechas pueden ser medidas en unidades arbitrarias, aunque pueden calibrarse en millones de años para cualquier gen dado, usando linajes donde el registro fósil es rico. Los puntos de bifurcación que antes hemos mencionado entre los linajes del ser humano y de los monos se han fechado a partir de pruebas de este tipo.
IV. PRUEBAS.
“En esta parte analizaremos seis de las maneras y técnicas que ha usado la comunidad científica para corroborar las teorías de la evolución. También demuestra un interés por darle un criterio de emparentamiento a las especies”
1. Paleontología:
La paleontología o registro fósil es el estudio de la vida prehistórica animal y vegetal, que realiza mediante el análisis de restos fósiles. El estudio de estos permite a los científicos determinar la historia de la evolución de organismos extintos, de la misma manera que si fueran organismos vivos. El estudio de los restos fósiles no sólo ofrece argumentos a favor del cambio y modificación (evolución) sufrida por antecesores de las especies actuales, sino que indica también que el proceso ha sido muy lento. El término fósil se refiere no solamente a las partes del cuerpo de un organismo que pueden preservarse, sino también a cualquier impresión o huella dejada por organismos antiguos.
En muchos casos se ha reconstruido el registro fósil completo de algún animal. Es el caso del caballo. El registro conocido comienza con Hyracotherium, del tamaño de un perro, con varios dedos en cada pata y dentición para ramonear, que aparece hace 50 millones de años, y finaliza con Equus, el caballo actual, mucho más grande, con sólo un dedo por pata y con dentadura apropiada para pastar. Se conservan muchas formas intermedias, así como otras formas que evolucionaron hacia otras ramas que no han dejado descendientes actuales. Otro ejemplo, es el de la mandíbula de los reptiles. Está formada por varios huesos; la de los mamíferos es de una sola pieza; los otros huesos de la mandíbula de los reptiles evolucionaron hasta convertirse en los que ahora forman parte del oído de los mamíferos. Esto puede parecer inverosímil, ya que es difícil imaginar las funciones intermedias de estos huesos. En respuesta a esto, se han descubierto dos tipos de terápsido (reptil de forma parecida a la de los mamíferos actuales) con una doble articulación mandibular: una compuesta de los huesos que persisten en la mandíbula mamífera y la otra por los huesos cuadrados. Eventualmente se dice que esto dio lugar al martillo y al yunque del oído de los mamíferos. Estas formas intermedias son las que nos permiten ver o analizar la transición de una especie a la otra.
Los fósiles constituyen un registro de animales y plantas que vivieron en épocas anteriores, el cual nos permite comprender dónde y cuándo vivieron y nos da una idea de los tipos de ambiente en que habitaron. Cuando se han encontrado suficientes fósiles de organismos de diferentes edades geológicas, es posible rastrear las líneas de evolución que los originaron.
2. Pruebas Anatómicas:
Al comparar dos organismo en su anatomía se pueden establecer un sinnúmero de comparaciones. Esta rama de la ciencia también nos permite deducir cambios evolutivos por ejemplo en comparaciones funcionales y estructurales, al observarse similitudes entre especies. Estas similitudes son llamadas analogías y homologías anatómicas. Las estructuras que tienen un origen común, pero no necesariamente conservan la misma función, se denominan homólogas y constituyen una evidencia a favor de la hipótesis de que estas seis especies derivan de un mismo ancestro común.
Un ejemplo clásico de homología lo constituye el miembro anterior de los tetrápodos. El ala de un ave, la aleta de una ballena, la pata de un caballo y el brazo de un hombre, a pesar de tener funciones distintas como volar, nadar, correr, o agarrar, comparten un mismo patrón estructural: todas estos miembros están formados por los mismos tipos de huesos (un húmero, un radio, un cúbito, una serie de metacarpales y, en términos generales, cinco dígitos). Los distintos huesos de las extremidades anteriores de los animales de la figura se muestran en color para indicar las similitudes fundamentales de estructura y organización. ver cuadro (5)
Mientras que la homología nos permite distinguir relaciones de descendencia, las analogías son un problema al momento de reconocer similitudes compartidas por una historia evolutiva en común. Ejemplos de órganos análogos son aquellos que desempeñan la función por ejemplo de volar en organismos tan distintos como un pterodáctilo (reptil volador de la era secundaria), una ave, una murciélago y un insecto. El origen del ala en cada uno de ellos es distinto: el los insectos es una membrana sostenida por venas quitinosas, en las aves son las plumas junto con la estructura ósea, en el murciélago es un pliegue de la piel que se sustenta fundamentalmente por el alargamiento de cuatro de los dedos, mientras que en el pterodáctilo el pliegue cutáneo su sustenta por el alargamiento de un solo dedo.
Existe una tercera clasificación de los órganos, son los órganos vestigiales o rudimentarios, se trata de aquellos órganos que en algunas especies no desempeñan función alguna. esto se explica por la postulación de Lamarck, que dice que un órgano o estructura puede desarrollarse o desaparecer según cuánto se utilice. Tal es el caso de la ballena, que presenta huesos homólogos a los huesos de la cadera de otros vertebrados. El ser humano también presenta órganos rudimentarios. El apéndice, por ejemplo, es vestigio de un órgano presente en conejos que se ocupa de almacenar celulosa para digerirla con bacterias. Esto es evidencia de nuestros antepasados herbívoros. Tal es el caso también de nuestras “muelas del juicio” y la “piel de gallina”.
Estas clasificaciones son comparables con dos mecanismos de evolución: divergente y convergente. La primera se da cuando una población se aísla del resto de la especie, y en el tiempo evoluciona de una manera diferente a la original. La forma que tomará la evolución del subgrupo dependerá del ambiente en que se desarrolle. Se afirma que el origen del oso polar es a causa de la evolución divergente. El registro fósil nos indica que en una de las glaciaciones del pleistoceno una población de oso pardo quedó aislada en el polo norte, y por la hostilidad del medio evolucionó en el actual oso polar. Este cambio es notable en la dieta de ambos, uno totalmente carnívoro y el otro herbívoro; la dentadura, su color, el pelaje y la forma de sus plantas, en el caso del oso polar tienen tracción y cubrimiento especial para poder desplazarse por la nieve.
El segundo tipo de evolución nombrado se da cuando los organismos evolucionan y se adaptan a ambientes similares, y aunque dichos organismos tengan una relación muy lejana, suelen parecerse entre sí. Es el caso de la ballena y el tiburón; la ballena es un mamífero con antepasados terrestres, por lo que posee pulmones, y sangre caliente, y la estructura ósea de sus aletas son homólogas a una mano de su ancestro tetrápodo y en cambio el tiburón es un pez y posee branquias. Esto nos demuestra que a pesar de tener un origen muy distante, y una estructura distinta, presentan rasgos externos muy similares, dada su adaptación al medio acuático.
Existe un tercer tipo de evolución; llamada evolución paralela, empleada en una situación en la cual los linajes han mutado de maneras similares, de modo que sus descendientes se parecen tanto como lo fueron sus antepasados, como es el caso de la evolución de los marsupiales en Australia, como el koala y el canguro.
Es posible establecer una relación entre los órganos homólogos y análogos, con la evolución divergente y convergente, ya que la evolución divergente lleva a la formación de órganos homólogos, pues al aislarse poblaciones de una especie, deben mutar para adaptarse a su nuevo medio y de esta forma conservan su estructura base, pero la función de sus partes puede llegar a ser totalmente distinta. En cambio, la evolución divergente forma órganos análogos, pues el tener que adaptarse a un mismo medio, lleva al desarrollo de estructuras similares, pues las condiciones externas son las mismas o muy parecidas.
3. Pruebas Embriológicas:
Es impresionante el parecido del desarrollo embrionario de todos los vertebrados, especialmente en sus primeras etapas (segmentación y morfogénesis). Se podría decir, que estas similitudes son lógicas, ya que todo vertebrado tendría que partir con las mismas características, pero esto no lo explica todo. Hay estructuras de vertebrados de organización avanzada que aparecen durante el desarrollo, estas son típicas de vertebrados más primitivos, pero aparecen o se modifican hasta que son irreconocibles. Por ejemplo el embrión humano en un principio posee una cola que al avanzar el desarrollo desaparece, es el mismo caso de un corazón con dos cámaras que por ejemplo en el ser humano se transforman en cuatro, pero en otros vertebrados toman rumbos distintos.
Estos cambios no sólo son similares entre los vertebrados actuales, sino que también se asemejan mucho al desarrollo de los vertebrados primitivos; a esto se le llama recapitulación y nos es posible conocerla gracias al registro fósil. La recapitulación anatómica puede ir acompañada de una recapitulación bioquímica, esto se explica por ejemplo, con los desechos que excretan los peces, que es el mismo que excretan los renacuajos hasta que experimenta metamorfosis y se transforman en ranas adultas, ya que el desarrollo embrionario de peces y ranas es muy similar en etapas tempranas. De estas pruebas de se deduce que cuanto más distante sea la relación entre dos vertebrados más corto será el período durante el cual pasen por estados de desarrollo similares.
4. Parentescos bioquímicos:
El estudio de la anatomía comparada demuestra la presencia de homologías anatómicas así como el estudio bioquímico de diferentes organismos ha revelado homologías bioquímicas. De esta forma, se han observado enzimas citocrómicas que se hayan en casi todos los organismos vivos. Una de estas , el citocromo c es una cadena polipéptica que consta de 104 a 112 a.á. Se ha podido determinar la secuencia exacta de estas cadenas en organismos tan diversos como desde el hombre hasta el moho de pan (neurospora). Aunque hay variación considerable de las secuencias en especial entre organismos relacionados remotamente, también existe semejanza considerable. Por ejemplo, la secuencia del hombre difiere de la secuencia del mono rhesus solo en un sitio de la cadena, mientras que el citocromo c de la planta de trigo presenta 35 diferencias. No obstante los demás 35 aminoácidos en las respectivas cadenas, son los mismos en las especies estudiadas. Un intervalo de la cadena que comprende 11 aminoácidos de la cadena, son comunes a todos los organismos conocidos. Seguramente este fenómeno significa que todos heredamos este gen de un mismo antecesor común, aunque modificado por una acumulación de mutaciones. ver cuadro (2)
Por otro lado, la mayoría de los vertebrados poseen hormonas similares. Por ejemplo la prolactina está presente en los peces, en las aves y en los mamíferos, aunque su función es distinta en todos ellos. Por lo que es correcto afirmar que es una hormona heredada a partir de un antecesor común aunque con función modificada según la forma de vida de cada animal.
Existe también un método llamado serología comparada, que se explica de la siguiente manera: si mezclamos en un tubo de ensayo suero de sangre de conejo con anticuerpos antihumanos con suero humano, se forman complejos insolubles antígeno-anticuerpo. Estos anticuerpos, reaccionan también con suero sanguíneo de otros mamíferos, pero en distinta cantidad. Entonces, si mezclamos los anticuerpos antihumanos, con suero de un hombre, con un antropoide, un mono del viejo mundo y otro del nuevo mundo, de un cerdo y de otros animales (por separado) por ejemplo, se producirá el precipitado antígeno-anticuerpo; pero la cantidad disminuirá desde el hombre hasta el cerdo en este caso. La cantidad de suero corresponde estrechamente con el grado de parentesco actualmente aceptado entre el hombre y los demás mamíferos. Mientras más relacionados estén, mayor será la cantidad del complejo antígeno anticuerpo, porque si dos especies se separaron tardíamente en la escala evolutiva, sus mecanismos bioquímicos de defensa pueden ser similares. ver cuadro (3)
De estos tres casos anteriores se deduce que el grado de similitud estructural u homología bioquímica, es aproximadamente proporcional a la cercanía del parentesco comparado. La bioquímica es una fuente de comparación mucho más fidedigna, ya que se pueden establecer relaciones fáciles de medir como la cantidad de a.á. distintos o la cantidad de una sustancia, pero existen infinidades de criterios a comparar.
5. Aislamiento Geográfico:
Una manera de explicar el origen de las especies, es por la situación geográfica. Esto plantea que si una población de una especie, queda separada del grupo por una barrera geográfica, es posible, que la parte separada (al igual que el resto) evolucionen por cualquiera de los mecanismos nombrados, pudiendo variar así, sus costumbres, hábitat, dieta alimenticia, instintos, etc. Si ambos grupos difieren lo suficiente el uno del otro, van a llegar a un punto en el que se produzca especiación. Y si eso sucede, aunque la barrera geográfica fuera superada o desapareciese, los individuos de ambas especies no podrían intercambiar genes. Un ejemplo es el ya citado anteriormente de la divergencia del oso polar a partir del osos pardo. Este proceso puede explicarse como parte de la microevolución, en la que juega un rol muy importante. ver cuadro (4)
6. Etología:
La etología es el estudio del comportamiento animal. Este estudio también nos da pruebas para demostrar cuan emparentadas están las especies, según los hábitos e instintos que tienen. Por ejemplo las varias especies de patos que existen se comunican por medio del lenguaje visual en el cortejo. Hembra y macho realizan movimientos que son estímulo para que el contrario realice el siguiente. En la actualidad, existen muchas especies de estas aves, pero cada una realiza una “danza” de cortejo diferente. Entonces, se dice que mientras más similares sean estos movimientos, o más movimientos en común tengan, más cercanas serán ambas especies estudiadas. Si divergieron de un ancestro común, en algún momento de su escala evolutiva habrán tenido el mismo “rito sexual”, los cuales fueron cambiando, produciendo así un aislamiento sexual. Este aislamiento hace que no haya más intercambio de genes entre las especies de pato, perpetuando la especie como tal.
V CONCLUSION - OPINIÓN GRUPAL.
Después de analizar pruebas y teorías, y comprender más de lo que significa la evolución, mutación y otros términos y conceptos, sin necesariamente entender las propiedades de la genética, nos damos cuenta de que a pesar de que no podemos observar la evolución como un todo, se pueden recurrir a otras ingeniosas técnicas de medición de la evolución y reconocimiento de sus mecanismos. El análisis de los mecanismos evolutivos requiere se un estudio muy complejo y multidisciplinario, por lo que reúne a paleontólogos, bioquímicos, anatomistas, genéticos, botánicos, sólo por mencionar las pruebas mostradas a través de esta investigación.
Además, nos lleva a proyectarnos hacia el futuro, ya que esta rama de la biología ha tomado mucha fuerza en sólo un siglo, y somos partidarios de que el hombre logrará en un futuro cercano comprender los complejos avances evolutivos y todo lo relacionado a ellos a pesar de que por momentos el problema pareciera que está fuera de los límites de la ciencia como en el caso de Darwin y las mutaciones. Quizás no estamos al alcance aún de encontrar las respuestas.
Creemos también que si se descubren las respuestas algún día más que una revolución científica, será una revolución religiosa, donde todo Credo podría tomar un curso diferente al descartarse el creacionismo “totalmente”. No solamente porque los Credos apoyan el creacionismo, sino que también porque el hombre será creador.
Pero remontándonos al futuro: ¿qué pasaría si el ser humano logra dominar la técnica de la evolución? ¿Seremos capaces de crear ya no sólo líneas genéticas superiores de pollos, sino que una nueva especie donde sólo importe el aumento del peso cárnico dejando de lado todo elemento y equilibrio ecológico? ¿Seremos capaces de pensar antes en todas las repercusiones que puede tener esto sobre el equilibrio natural y sobre el ser humano mismo? ¿qué pasaría si se aplicara esta tecnología a los seres humanos ?¿evolucionaríamos sin límite? ¿tendrán solo algunos acceso a esta información y lo que decide la naturaleza bajo selección natural lo decidirán un par de personas acerca de quiénes son los más aptos para evolucionar? En este sentido al ser humano todavía le falta por evolucionar y no estamos listos aún para tan preciado misterio.
BIBLIOGRAFÍA.
BIOLOGÍA; HELENA CURTIS. Editorial Panamericana, 4ª Edición, Buenos Aires, 1990.
BIOLOGÍA; HELENA CURTIS, N. SUE BARNES. Editorial Panamericana, 6ª Edición, España, Septiembre 2000 (libro y CD).
BIOLOGÍA; JOHN W. KIMBALL. Editorial Addison-Wesley Iberoamericana, 4ª Edición, E.U.A., 1986.
BIOLOGÍA; NATALIO GLAVIC, GRACIELA FERRADA. Ediciones Pedagógicas Chilenas, Chile 1990.
ENCICLOPEDIA MICROSOFT ENCARTA; MICROSOFT CORPORATION. E.U.A., 1998.
VI. ANEXO(s)
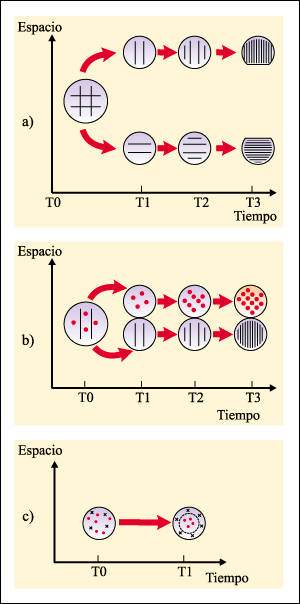
(1)Especiación
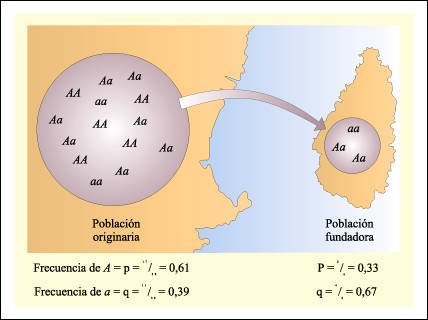
(4) Aislamiento geográfico.
(2) Los números indican la cantidad de aminoácidos por los cuales cada citocromo c difiere del citocromo c correspondiente al punto de bifurcación más cercano. Las líneas de guiones indican que se ha acortado una rama y que, por lo tanto, no está representada a escala.
(3) Tabla de reacción entre anticuerpos antihumanos (preparados en el conejo) y sueros de varios mamíferos tomados del humano como 100%
| Humano | 100% |
| Chimpancé | 97% |
| Gorila | 92% |
| Gibón | 79% |
| Babuino | 75% |
| Mono Araña (especie del nuevo mundo) | 58% |
| Lemur | 37% |
| Erizo (insectívoro) | 17% |
| Cerdo | 8% |
ÍNDICE.
Portada 1
Introducción. 2
Definición de Concepto. 3
Teorías de la Evolución.
Svante Arrhenius 3
George Cuvier 4
Jean-Baptiste de Lamarck 4
Charles Darwin y Alfred Russel Wallace 5
Hugo de Vries 8
Teoría Sintética de Evolución 8
Teoría Neutralista 11
Evidencias de la Evolución
Paleontología 12
Pruebas Anatómicas 13
Desarrollo embrionario 15
Parentescos bioquímicos 15
Aislamiento geográfico 16
Etología 17
Conclusión- Opinión Grupal 18
Bibliografía 19
Anexo(s) 20
Descargar
| Enviado por: | José Manuel Nano Campos |
| Idioma: | castellano |
| País: | Chile |
Todos los derechos reservados.