Biología, Botánica, Genética y Zoología
Energía de los organismos vivos
REQUERIMIENTOS DE ENERGÍA DE LOS ORGANISMOS VIVOS
Los organismos vivos pueden clasificarse en dos grandes grupos: autótrofos y heterótrofos.
Los heterótrofos, incluyen a todos los animales, hongos y muchos microorganismos, son aquellos organismos que dependen también del consumo continuo de una gran variedad de compuestos orgánicos que toman del medio ambiente. Estos compuestos aportan el esqueleto de átomos de carbono que necesitan para sus procesos biosintéticos y, mediante la oxidación controlada de los mismos, la energía metabólica requerida para todos los procesos celulares.
Los autótrofos son aquellos organismos que poseen la capacidad de sintetizar sus propias sustancias complejas a partir de elementos simples como el CO2 y el H2O, utilizando para ello la energía proveniente de distintas fuentes. De acuerdo con la fuente de energía que utilicen, los organismos autótrofos pueden clasificarse en:
1) autótrofos quimiosintéticos, que utilizan la energía proveniente de reacciones de oxidorreducción, y
2) autótrofos fotosintéticos, que emplean como fuente de energía la luz solar. En el grupo de los organismos autotróficos, los fotosintéticos constituyen el grupo más importante.
Se cree que todos los organismos heterotróficos primitivos de nuestro planeta tuvieron acceso a grandes cantidades de compuestos orgánicos formados en procesos geoquímicos. Sin embargo, esos compuestos se agotaron hace miles de millones de años y desde entonces los compuestos orgánicos requeridos por los heterótrofos son producidos por los autótrofos, especialmente los fotosintéticos. Es por ello que a los organismos autótrofos, y en particular a los fotosintéticos, les corresponde la importante función de restaurar el equilibrio, consumiendo CO2, que transforman en sustancias orgánicas, y generando simultáneamente oxígeno, que es empleado por los heterótrofos en las reacciones oxidativas de degradación. Es decir, los organismos fotosintéticos mantienen en la biosfera el equilibrio de compuestos esenciales, como el CO2 y el 02, y al mismo tiempo, merced de la síntesis de compuestos orgánicos, mantienen a los heterótrofos.
Definición y ecuaciones del proceso fotosintético:
De todas las formas en que la luz interacciona con la materia viva, la más importante es la fotosíntesis, que ya hemos definido como el proceso en virtud del cual la energía electromagnética es captada por ciertos organismos especializados y convertida en energía química.
Una gran variedad de organismos y, entre ellos, los procariotes eucariotes son capaces de realizar fotosíntesis. En el grupo de los eucariotes, las plantas verdes, las algas multicelulares y unicelulares y ciertos organismos unicelulares como los euglenoides, dinoflagelados y las diatomeas poseen esa capacidad. Los procariotes fotosintéticos comprenden las algas verdeazuladas y ciertas bacterias de los géneros Chlorobium, Chromatium y Rhodospirillum.
La reacción global de la fotosíntesis en plantas superiores pueden expresarse en una sencilla ecuación:
![]()
donde ![]()
es la fuente de carbono y el agua el reductor que aporta los átomos de hidrógeno necesarios para la biosíntesis de carbohidratos, obteniéndose además oxígeno como subproducto. Los productos de la reacción tienen mayor energía libre que los reactantes y, por lo tanto, es posible observar que la energía del sol es captada parcialmente por los organismos fotosintéticos en la glucosa y el oxígeno que se producen.
Al analizar el proceso fotosintético conviene dividirlo en dos grandes etapas, o fases, conocidas como reacciones luminosas y reacciones oscuras de la fotosíntesis. En las reacciones dependientes de la luz, que definen a la fase luminosa, la energía electromagnética es captada por un sistema especializado de pigmentos y utilizada para sintetizar ATP y NADPH (nicotinamida adenindinucleótido fosfato reducido).
En las reacciones oscuras --fase oscura de la fotosíntesis-- el ATP y el NADPH formados en la fase luminosa se utilizan como fuente química de energía y poder reductor, respectivamente, para convertir el CO2 en carbohidratos. Estas reacciones comienzan en el estroma del cloroplasto y continúan en el citoplasma de la célula, denominándose reacciones oscuras porque, si bien utilizan los productos de la fase luminosa, no requieren directamente luz.
El Cloroplasto y su organización
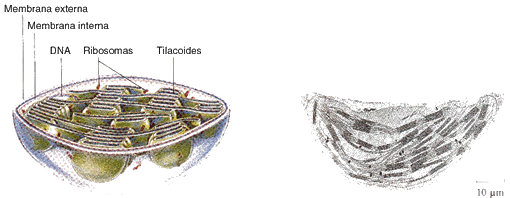
En las células eucariotes, el proceso fotosintético se encuentra localizado en un orgánulo subcelular membranoso que es característico de las células vegetales: el cloroplasto. Tanto la forma que adopta este orgánulo, como el tamaño y el número de ellos por célula, varía de una especie vegetal a otra.
Los cloroplastos están separados del citoplasma por una envoltura continua constituida por una doble membrana interna, cuya permeabilidad es selectiva y dentro de la cual se encuentran ciertas proteínas transportadoras. Entre los transportadores, el más importante es el translocador de fosfato, que constituye casi un 20% de las proteínas de la membrana interna y que participa en el transporte activo de las triosas fosfato. Se han descrito asimismo sistemas para el transporte activo de glucosa, piruvato, aminoácidos, ácidos dicarboxílicos y un intercambiador ADP-ATP. Esta membrana participa, además, en el proceso de importación de proteínas cloroplásticas codificadas por el ácido desoxirribonucleico nuclear (ADN* nuclear) y posee una ATPasa dependiente de Mg2+, cuya función fisiológica no se conoce aún.
La membrana interna rodea a un gran espacio central, llamado estroma, que contiene numerosos sistemas enzimáticos, donde tiene lugar la fijación de dióxido de carbono. Hay otras vías metabólicas que intervienen en la fijación de nitrógeno y de aminoácidos. En dicho espacio funciona un sistema genético completo diferente del nuclear que se encarga de la síntesis de unos cien polopéptidos cloroplásticos.
En el interior de los cloroplastos se observa un tercer tipo de estructura membranosa dispuesta en forma de sacos aplastados, o discos, llamados tilacoides. Los tilacoides se apilan y constituyen estructuras mayores, denominadas granas. El espacio interior, o lumen, de cada tilacoide se conecta a veces con el lumen de otros tilacoides, formándose de este modo un tercer compartimento interno, o espacio intratilacoidal, que está separado del estroma por la llamada membrana tilacoidad.
En los tilacoides tienen lugar las reacciones de la fase luminosa de la fotosíntesis. En estas membranas la relación proteína-lípido es elevada, abundan los lípidos neutros, en especial galactolípidos, y se encuentran los pigmentos que captan la energía solar, así como los complejos proteicos asociados a la clorofila en los centros de reacción, los componentes de la cadena transportadora de electrones que participan en la obtención de NADPH y los del complejo ATPasa capaz de catalizar la formación de ATP a partir de ADP (adenosina-difosfato) y pi (ortofosfato inorgánico).
Aspectos Metabólicos del Cloroplasto
El ciclo reductivo de las pentosas fosfato, más conocido como ciclo de Calvin y Benson, ocupa una posición central en el metabolismo de los cloroplastos. Este ciclo utiliza al ATP y NADPH producidos durante la fase lumínica de la fotosíntesis para lograr la asimilación reductiva del CO2 hasta convertirlo en carbohidratos. A continuación, los carbohidratos así formados son utilizados en el interior de los cloroplastos como precursores para la síntesis de almidón y en el citoplasma de la célula para sintetizar sacarosa.
Fuente: Andreo y Vallejos, Fotosíntesis, monografía de la OEA.
Preguntas Frecuentes:
-
¿ Cuál es la importancia de los organismos autotrofos para la vida en el planeta?
-
Establezca una comparación entre organismos autotrofos y organismos heterotrofos
-
Explique la organización del cloroplasto.
-
Escriba la ecuación general de la fotosíntesis
-
Establezca una comparación entre fase clara y fase oscura
-
Señale la importancia de los pigmentos fotosintéticos.
REQUERIMIENTOS DE ENERGÍA DE LOS ORGANISMOS VIVOS
FOTOSINTESIS: FASE CLARA O FOTOLISIS DEL AGUA
Los pigmentos fotosintéticos
En el proceso fotosintético participan tres tipos de pigmentos distintos: clorofilas, carotenoides y ficobilinas. La función de estos pigmentos consiste en proporcionar a los organismos fotosintéticos un sistema eficiente para la absorción de la energía electromagnética. Esta energía es posteriormente transferida hacia los centros de reacción donde ocurren las reacciones fotoquímicas primarias.
En las plantas superiores y en las algas verdes --organismos fotosintéticos que generan oxígeno la clorofila es de dos tipos: clorofila a y clorofila b; en cambio, las bacterias fotosintéticas que generan oxígeno poseen sólo bacterioclorofila b.
La clorofila a es un complejo magnesio-porfirínico compuesto por un tetrapirrol cíclico (I a IV) que posee un anillo de cíclopentanora (V) fusionado. Los cuatro átomos de N de los anillos pirríticos están coordinados con un átomo de magnesio formando un complejo glanar estable. La molécula de clorofila posee, además, una cadena terpenoide constituida por el alcohol fitol, que se encuentra esterificando a un residuo de propionato, sustituyente de uno de los anillos pirrilicos (IV). Este alcohol de cadena larga, compuesto de cuatro unidades de isopreno, confiere a la molécula de clorofila la característica de ser altamente hidrofóbica. La clorofila b difiere de la clorofila a en que contiene como sustituyente un grupo formilo (-CHO) en vez de mitilo (-CH3) en el anillo pirrólico II.
La gran eficiencia de las clorofilas como fotorreceptores se correlaciona con la presencia alternante de enlaces simples y dobles en su estructura. Las clorofilas presentan máximos de absorción en la región visible del espectro, pero difieren en sus espectros de absorción. Por ejemplo, la luz de 460 nm no es apreciablemente absorbida por la clorofila a mientras que sí puede ser captada por la clorofila b, complementándose de este modo la absorción de la luz incidente. En la región del espectro comprendida entre 500 y 600 nm la absorción de la luz por las clorofilas es muy débil.
A pesar de que las clorofilas son cuantitativamente las moléculas fotorreceptoras que predominan en las plantas y algas verdes, existen también pigmentos accesorios que presentan máximos de absorción a longitudes de onda distintas de las clorofilas y que actúan como moléculas receptoras de luz suplementarias en regiones del espectro en que las clorofilas no tienen absorción, aumentando así la eficiencia con que las células fotosintéticas captan la luz.
-
Los pigmentos accesorios más importantes: son los carotenos, siendo el carotenoide el más frecuente, especialmente en las plantas verdes. Los carotenos son largas cadenas poliisoprenoides con dobles enlaces conjugados, que poseen en ambos extremos anillos de ciclohexano sustituidos. Los cloroplastos también contienen una gran variedad de carotenoles que son carotenoides oxigenados, denominados comúnmente xantofilinas. Entre estas, la neoxantina, violaxantina y luteína predominan en las plantas y algas verdes.
-
Una tercera clase de pigmentos accesorios, la ficobilinas, aunque de distribución más limitada, están presentes tanto en las algas rojas como en las algas verdeazuladas y se caracterizan estructuralmente por ser tetrapirroles de cadena abierta. Las ficocianinas, proteínas conjugadas que poseen ficocianobilina como pígmento unido, confieren el color característico a las algas verdeazuladas. Del mismo modo, las ficoeritrinas, proteínas conjugadas que contienen ficoeritrobilina, predominan en las algas rojas y dan a estas algas su color característico.
|
|
|
|
Organización de los Pigmentos Fotosintéticos
A los pigmentos fotosintéticos les corresponde la función de captar la energía electromagnética y transferirla eficientemente hacia el centro o los centros donde ocurre la reacción fotoquímica primaria. Para cumplir esta función las moléculas fotorreceptoras forman agregados multimoleculares compuestos por cientos de moléculas que se mantienen firmemente asociadas a la membrana tilacoidal por la acción de ciertas proteínas especiales. En estos agregados multicelulares, o unidades fotosintéticas, denominados fotosistemas, las proteínas especiales alteran a las moléculas fotorreceptoras de forma tal que la energía luminosa puede ser captada con igual eficiencia por cualquiera de ellas, para ser luego conducida hacia una molécula especial de clorofila llamada "clorofila centro de reacción". Cada fotosistema actúa, pues, como un embudo que recoge la energía luminosa y la conduce hacia un simple centro de reacción.
En las plantas superiores se distinguen dos tipos de fotosistemas: el fotosistema I y el fotosistema II. El primero está compuesto por unas 250 a 300 moléculas de clorofila a (entre las cuales predominan las que absorben a longitudes de onda más largas), una reducida proporción de clorofila b y carotenoides. Asociado al fotosistema I se encuentra una molécula de un pigmento fotooxidable especializado, denominado P 700, que constituye el centro de reacción de este fotosistema.
El fotosistema II contiene clorofila a (entre las cuales predominan las que absorben a longitudes de onda mas cortas), una proporción relativamente elevada de clorofila b y xantofilinas. El centro de reacción de este fotosistema es un pigmento fotooxidable llamado P680.
La energía lumínica que puede ser absorbida por una molécula aislada de clorofila en disolución se libera rápidamente como luz (fluorescencia) y calor al retornar la molécula a su nivel energético inicial.
En cambio, en un fotosistema, la molécula de clorofila centro de reacción se encuentra asociada en alto grado a un dador y a un aceptor de electrones y, merced a una reacción mediada por las proteínas especiales, el electrón excitado en el centro de reacción es transferido al aceptor de electrones dejando un hueco cargado positivamente con alta afinidad por los electrones. Este hueco es rápidamente ocupado por un electrón proveniente del dador de electrones.
De este modo, la organización en fotosistemas permite que la luz haga posible la transferencia neta de un electrón a partir de un débil dador de electrones hacia una molécula que se convierte, como resultado de esa transferencia, en un fuerte dador de electrones. Es decir, la energía de excitación que se liberaría rápidamente como fluorecencia y calor en una molécula aislada de clorofila es utilizada por los fotosistemas par elevar la energía de un electrón y crear un fuerte dador de electrones.
La función del fotosistema II, presente en algas y plantas verdes, es la de generar poder oxidante como prerrequisito para producir oxígeno a partir del agua. Las bacterias fotosintéticas, que no emplean el agua como reductor y en las cuales no se produce oxígeno como consecuencia del proceso fotosintético carecen del fotosistema II y contienen exclusivamente el fotosistema I.
Fase Bioquímica (10-4 a 10-2 segundos)
Esta fase comprende, debido a la participación de enzimas y transportadores de electrones, la liberación de oxígeno, la formación de poder reductor bajo la forma de NADPH, la síntesis de ATP a partir de ADP y fosfato y, por último, la asimilación reductiva del CO2 que puede representarse por la ecuación siguiente:
CO2 + 2 NADPH + 3 ATP ---- (CH20) + 2 NADP+ 3 ADP +3H
Transporte Fotosintético de Electrones
Actualmente se acepta que el mejor modelo para la interpretación del transporte fotosintético de electrones en organismos cuyo proceso fotosintético involucre el desprendimiento de oxígeno, es aquel que incluye dos reacciones fotoquímicas que actúan cooperativamente en serie. El fotosistema II genera en la luz un reductor débil y un oxidante fuerte capaz de reaccionar con agua para generar oxígeno mediante la reacción:
H2O-------------- 2 H+ + 2e- + 1/2 O2
Simultáneamente, el fotosistema I produce en la luz un oxidante débil y un reductor fuerte que puede reducir al NADP+. Los electrones tomados del agua por el fotosistema II son conducidos hacia el fotosistema I mediante reacciones de oxidorreducción que tienen lugar entre los dadores y aceptores de electrones localizados asimétricamente en la membrana tilacoidal que constituyen la cadena fotosintetica de transporte de electrones.
El transporte de electrones entre el reductor débil generado por el fotosistema II y el oxidante débil producido por el fotostema I es un proceso independiente de la luz.
El transporte de electrones desde el agua, que haría las veces de reductor, hasta el NADP+, que actuaría como oxidante, es unidireccional, por lo que se le denomina transporte de electrones "lineal" o "no cíclico", y puede ser representado por la ecuación:
H2O + NADP+---hv----1/2 O2 + NADPH + H+
El flujo de electrones se iniciaría con la absorción de un cuanto por uno de los fotosistemas. De este modo, la absorción de un cuanto por el fotosistema II resultaría en la eliminación de un electrón del agua y en la reducción del aceptor primario x 320. Los electrones del compuesto x 320 trfuvifo don transportados, entonces, mediante una serie de reacciones de oxidorreducción al pigmento P700. La absorción de un segundo cuanto de luz por el fotosistema I resulta en la oxidación del del P700 y en la reducción del P430, denominado aceptor primario de electrones del fotosistema I. Finalmente, el P430 reducido reduce al NADP+ merced a dos intermediarios proteicos.
Con respecto a la cadena de transportadores que conecta a ambos fotosistemas procede señalar que varios de sus componentes han sido aislados y caracterizados. El transporte fotosintético de electrones ha sido estudiado fundamentalmente en cloroplastos rotos, y el conocimiento que se tiene del mismo ha sido logrado gracias al empleo de aceptores
Aceptores de Electrones del Fotosistema
Actualmente se considera como aceptor primario de electrones del fotosistema I a un complejo en el que la ferredoxina está fuertemente asociada con una sulfoferroproteína. Este complejo se denomina P430 por presentar un máximo de absorción a 430 nm, que es inducido por iluminación de los cloroplastos. El mecanismo de reacción entre el complejo P430, que contiene ferredoxina unida, y la ferredoxina soluble aún no se conoce.
La ferredoxina, una ferroproteía no hemínica que actúa como aceptor secundario de electrones del fotosistema I, es el único transportador soluble de la cadena lineal del transporte fotosintético de electrones.
La reducción del NADP+, aceptor terminal de electrones, por la ferredoxina es catalizada por la enzima ferredoxina-NADP+ oxidorreductasa. Esta enzima fue descubierta por su actividad como diaforasa y tranhidrogenasa, y su participación en la reducción del NADP+ se inhibía por anticuerpos preparados contra la enzima.
La naturaleza hidrofílica de los aceptores artificiales de electrones que son reducidos por el fotosistema I (ferricianuro, sales de tetrazolio, quinonas y sales de dipiridilo) y los resultados de estudios realizados con anticuerpos e inhibidores indican que el lado aceptor del fotosistema I se orienta en la membrana tilacoidal hacia la superficie externa. Es decir, el sitio aceptor de electrones del fotosistema I, que comprende a la ferredoxina, la ferredoxina-NADP+ oxidorreductasa y a un componente estructural, estaría expuesto al medio hidrofílico del estroma, donde se encuentran las enzimas responsables de la asimilación oscura del CO2.
Dadores de Electrones del Fotosistema I
El descubrimiento de inhibidores de la plastocianina, como histonas, polilisinas y KCN, que inhiben el transporte de electrones entre el citocromo f y el fotosistema I, permitió considerar a la plastocianina y no al citocromo f como dador endógeno de electrones de este fotosistema.
La plastocianina es una cuproproteína localizada en la cara interna de la membrana tilacoidal, cuyo peso molecular es 10.000.
El citocromo, una hemoproteína firmemente unida a la membrana tilacoidal, está localizado en la ruta principal del transporte de electrones y actúa como reductor de la plastocianina y como oxidante de la plastoquinona. Recientemente se ha logrado purificar un complejo compuesto por citocrono b6, citocromo f y un centro hierro-azufre que exhibe la actividad plastoquinolplastocianina oxidorreductasa.
El empleo de anticuerpos específicos contra plastocianina y citocromo f, como asimismo estudios sobre dadores artificiales de electrones para el fotosistema I que poseen distinta lipofilicidad y polaridad (indofenoles y p-finilendiaminas sustituidas), han sugerido que el sitio dador de electrones del fotostema I estaría localizado en la cara interna de la membrana tilacoide.
Aceptores de Electrones del Fotosistema II
No existen hasta la fecha pruebas directas acerca de la naturaleza química del aceptor endógeno primario (X320) del fotosistema II. En estudios acerca de los cambios de absorbencia que presentan las membranas tilacoidales al ser excitadas por iluminación durante breves periodos de tiempo en la zona del espectro próxima al ultravioleta, se ha sugerido que se trataría de una molécula de plastoquinona firmemente unida a la membrana que sólo podría ser reducida hasta el nivel de semiquinona.
Otro compuesto denominado C550, identificado como un betacaroteno, podría estar unido al centro de reacción del fotosistema II y actuaría como pigmento indicador del estado redox del aceptor primario de electrones de este fotosistema.
La plastoquinona se encuentra presente, en relación con los demás componentes de la cadena, en una proporción muy elevada --10 a 40 moléculas por centro de reacción-- y actuaría como aceptor secundario de electrones. La reducción de la plastoquinona tiene lugar en la cara externa de la membrana tilacoidal mientras que su reoxidación ocurre en la parte interna; el resultado es la translocación neta de dos protones desde el estroma hacia el interior del tilacolide.
Preguntas Frecuentes
-
Escriba las ecuaciones fotoquímicas de la fotolisis del agua.
-
Nombre los aceptores primarios de los electrones del fotosistema I
-
Nombre los aceptores primarios de los electrones del fotosistema II
-
Explique la importancia de la Plastoquinona.
-
Explique la arquitectura topográfica de la membrana tilacoidal.
Fuente: Monografía de la OEA, Fotosíntesis, Carlos Andreo y Rubén Vallejos
Descargar
| Enviado por: | El remitente no desea revelar su nombre |
| Idioma: | castellano |
| País: | Chile |
Todos los derechos reservados.