Biología, Botánica, Genética y Zoología
Comunicación intercelular
1. - INTRODUCIÓN.
El funcionamiento de todas las sociedades animales y humanas es posible gracias a la comunicación. Esta consiste en el intercambio de mensajes entre los individuos, (un individuo establece con otro un contacto que le permite transmitirle una información).
Fuera del mundo animal y humano también se producen fenómenos de comunicación. De este modo, algunos aparatos emiten mensajes a otros aparatos; así, un termostato "ordena" al sistema de calefacción que se ponga en marcha cuando baja la temperatura. Y en el interior de los organismos vivos, el cerebro recibe información y transmite instrucciones. Los biólogos han descubierto que la conservación y la reproducción de las células se basan en un intercambio de comunicaciones.
El cuerpo de un animal puede contemplarse como una sociedad o ecosistema, cuyos miembros son células, que se organizan en conjuntos cooperativos denominados tejidos, que, a su vez, se asocian formando grandes unidades funcionales denominados órganos. De este modo, podríamos asemejar una célula con un individuo, el cual establece una vida social, necesita comunicarse con otros y relacionarse con su ambiente, respondiendo a distintos estímulos. Así, para que una persona pueda pensar, actuar o, simplemente, existir, las células de su cuerpo deben comunicarse entre sí; comunicación que efectúan poniendo en marcha mensajeros químicos, como las hormonas circulantes y los neurotransmisores.
Estos datos permiten adivinar la importancia que posee el estudio de la comunicación intercelular.
Para que pueda establecerse "comunicación", deben intervenir en ella los siguientes factores:
a) El emisor, que produce o cifra el mensaje, y lo emite.
b) El receptor, que lo recibe y descifra.
c) El código: conjunto de signos que se combinan mediante ciertas reglas conocidas por el emisor y el receptor. (En el caso concreto de la comunicación intercelular, estaría constituido por aminoácidos, nucleótidos, etc).
d) El mensaje: contenido de las informaciones que el emisor envía al receptor. (En nuestro caso, señales químicas, eléctricas...)
e) El canal: vía por la cual circula el mensaje. (Así, la circulación sanguínea en la señalización endocrina; como se explicará posteriormente).
f) El contexto: situación en que se transmite el mensaje y que contribuye a su significado.
El circuito de la comunicación puede representarse con esta figura:
| MENSAJE | ||
| EMISOR | !CANAL! | RECEPTOR |
| CÓDIGO |
CONTEXTO
Con este estudio, pretendo poner de manifiesto la existencia de una "verdadera" comunicación a nivel celular. Entre otros objetivos, el trabajo propuesto persigue sentar los principios generales y la base molecular de la comunicación intercelular, así como los principales mecanismos implicados en ésta.
2.- PRINCIPIOS GENERALES DE LA COMUNICACIÓN INTERCELULAR.
La evolución de los organismos pluricelulares ha dependido de la capacidad de las células de comunicarse entre ellas. La comunicación entre las células es necesaria para poder regular su desarrollo y su organización tisular, para controlar su crecimiento y división y para coordinar sus diversas funciones.
La importancia y la complejidad de la comunicación intercelular en los animales superiores sugiere que una gran proporción de los genes de estos organismos está dedicada a estos procesos.
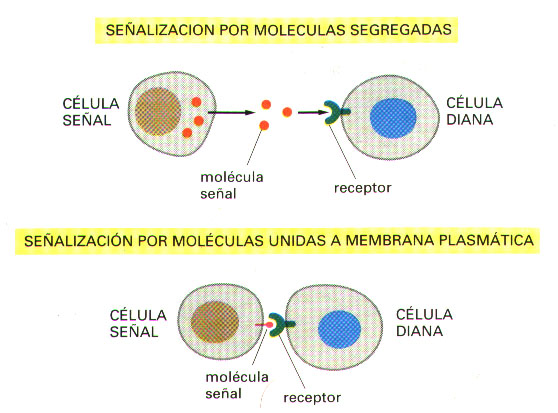
Las células animales se comunican de tres maneras: (Figura 1)
a) Segregan compuestos químicos que actúan como señales para células situadas a una cierta distancia.
b) Presentan moléculas señalizadoras, unidas a la membrana plasmática, que influyen sobre las células que establecen contacto físico directo con ellas.
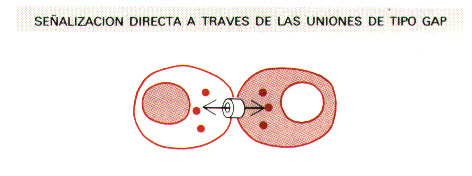
c) Forman uniones de tipo gap (uniones de tipo hendidura), que unen directamente los citoplasmas de las células que interaccionan, permitiendo así el intercambio de pequeñas moléculas.
Figura 1. Tres sistemas diferentes de comunicación de las células entre sí.
En general, podemos decir que las células de los animales superiores se comunican mediante centenares de tipos de moléculas señal, incluyendo proteínas, pequeños péptidos, aminoácidos, nucleótidos, esteroides, retinoides, derivados de ácidos grasos, e incluso gases disueltos como el óxido nítrico y el monóxido de carbono. Estas moléculas señal son liberadas por las denominadas células señal (emisoras del mensaje).
Sea cual sea la naturaleza de la molécula señal, la célula diana (receptora del mensaje) responde mediante una proteína específica denominada receptor. Éste se une específicamente a la molécula señal y, entonces, inicia una respuesta en la célula diana.
Muchas de las moléculas señal extracelulares actúan a concentraciones muy bajas y los receptores que las reconocen, usualmente, se unen a ellas con una elevada afinidad.
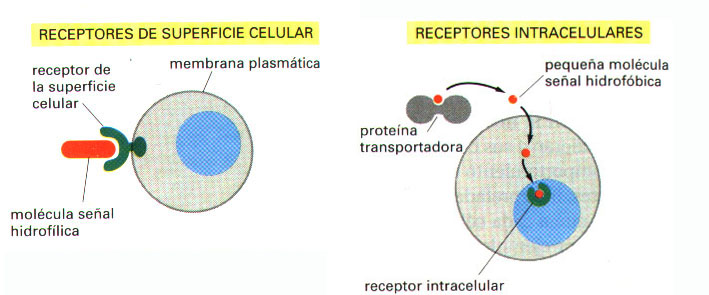
En la mayoría de los casos, los receptores son proteínas transmembrana de la superficie de las células diana; cuando se unen a una molécula señal extracelular (un ligando) se vuelven activos, de forma que generan una cascada de señales intracelulares (transducción de señales), que alteran el comportamiento de la célula. En algunos casos, sin embargo, los receptores están situados en el interior de la célula diana, y el ligando señal entra en la célula para activarlos: estas moléculas señal, por lo tanto, han de ser suficientemente pequeñas e hidrofóbicas para poder difundir a través de la membrana plasmática (Figura 2).
Figura 2. Las moléculas señal extracelulares se unen a receptores de la superficie celular o a receptores intracelulares. La mayoría de las moléculas señal son hidrofílicas, por lo que son incapaces de atravesar directamente la membrana plasmática; en lugar de ello, se unen a receptores de la superficie de la célula, los cuales, a su vez, generan una o varias señales en el interior de la célula diana. Por el contrario, algunas pequeñas moléculas señal difunden a través de la membrana plasmática y se unen a receptores situados en el interior de la célula diana -en el citosol o en el núcleo- (como se observa en la figura). Muchas de estas pequeñas moléculas señal son hidrofóbicas y casi insolubles en soluciones acuosas; por ello, son transportadas a través del torrente sanguíneo y de otros fluidos extracelulares, unidas a proteínas transportadoras, de las que han de disociarse antes de entrar en la célula diana.
En este trabajo, nos centraremos fundamentalmente en el estudio de la comunicación entre células animales mediada por señales químicas segregadas (comunicación a distancia). Este énfasis refleja el estado actual de conocimientos sobre el tema: las moléculas segregadas son mucho más fáciles de estudiar que las moléculas unidas a membrana, por lo que conocemos muchos más detalles de su actuación.
La comunicación o señalización dependiente de contacto, vía moléculas unidas a membrana, es mucho más difícil de estudiar y por ello mucho peor conocida; pero es de crucial importancia, especialmente, durante el desarrollo y en la respuesta inmune.
Primero, trataremos muy brevemente la comunicación que depende del contacto célula-célula a través de las uniones de tipo gap.
3. - UNIONES DE TIPO GAP.
Todas las células vivas están limitadas por una membrana plasmática, envoltura delgada que regula de manera selectiva el flujo de nutrientes y de iones que la atraviesa, del interior de la célula al medio externo o en dirección opuesta.
Una unión intercelular (o, simplemente, unión celular) es una región reducida de la membrana plasmática, que se ha especializado para establecer el contacto entre células. Las uniones intercelulares son esenciales para el desarrollo y el funcionamiento normal de todas las formas de vida superiores.
El tipo habitual de unión celular es la unión de tipo gap (gap junction), también denominada unión de tipo hendidura, que se encuentra en la mayoría de los tejidos de la práctica totalidad de las especies animales.
Las uniones de tipo gap median la comunicación intercelular al permitir el paso de iones inorgánicos y otras pequeñas moléculas hidrosolubles entre los respectivos citoplasmas, acoplando las células tanto eléctrica como metabólicamente. Este acoplamiento celular tiene importantes implicaciones funcionales.
Se trata de uniones especializadas célula-célula que pueden formarse entre membranas plasmáticas situadas en estrecho contacto, y que conectan directamente los citoplasmas de las células que unen, a través de estrechos canales llenos de agua. Los canales permiten el intercambio de pequeñas moléculas señal intracelulares (mediadores intracelulares), como el Ca 2+ y el AMP cíclico, pero no el de macromoléculas como proteínas y ácidos nucleicos. Así pues, las células conectadas por uniones comunicantes o de tipo gap pueden comunicarse entre ellas directamente, sin tener la dificultad de la barrera que supone la presencia de las membranas plasmáticas.
4. - SEÑALIZACIÓN QUÍMICA.
Los mecanismos de señalización química dependen de las distancias a las que transmiten la señal:
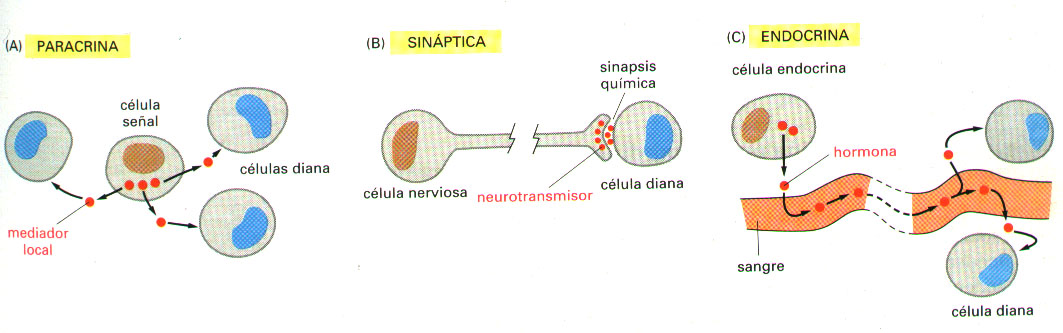
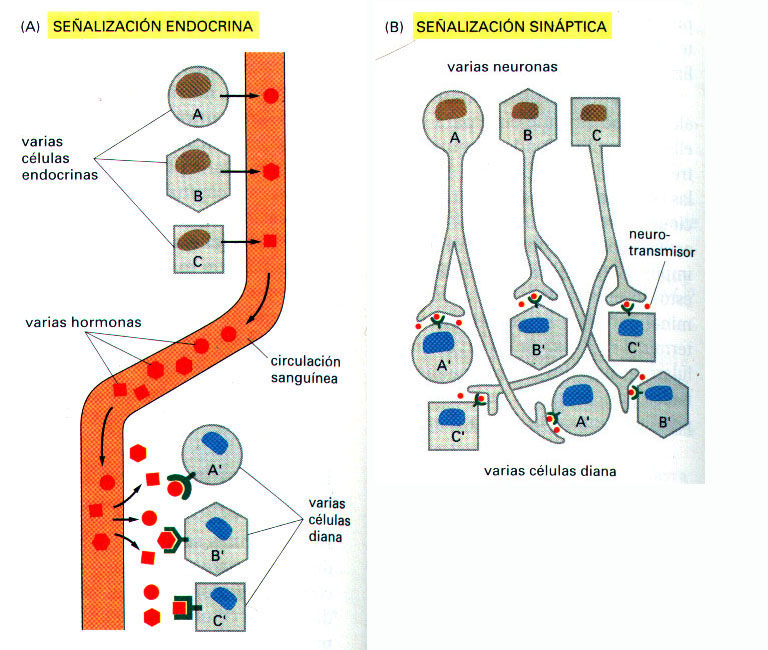
a) En la señalización endocrina, células endocrinas especializadas segregan sus moléculas señal, denominadas hormonas, las cuales viajan a través del torrente circulatorio (en un animal) o de la savia (en una planta), son transportadas a largas distancias y actúan sobre células diana que se hallan ampliamente distribuidas por todo el cuerpo. Las células endocrinas son células señal especializadas que controlan el comportamiento del organismo como un todo.
b) En la señalización paracrina, las células segregan moléculas señal que actúan como mediadores químicos locales, los cuales son tan rápidamente captados, destruidos o inmovilizados, que únicamente llegan a actuar sobre las células situadas en el entorno inmediato de la célula que los ha segregado (célula señal). De esta manera, las señales paracrinas solamente afectan a las células diana más cercanas.
c) En la señalización sináptica, exclusiva del sistema nervioso, las células segregan, a uniones especializadas llamadas sinapsis químicas, unos mediadores químicos de muy corto alcance denominados neurotransmisores, que sólo actúan sobre la célula diana inmediata (célula postsináptica adyacente). (Figura 3).
En cada caso, la célula diana responde a una señal extracelular particular mediante proteínas específicas (receptores), que unen la molécula señal e inician la respuesta. En las respuestas endocrina, paracrina y sináptica se utilizan muchas moléculas señal y muchos receptores comunes. Las diferencias cruciales entre estos sistemas de comunicación radican en la velocidad y en la selectividad con que las señales son transportadas hasta las células diana.
Figura 3. Tres tipos de señalización mediadas por moléculas segregadas. En las señalizaciones paracrina, sináptica y endocrina se utilizan muchos tipos iguales de moléculas señal.
Para un organismo pluricelular, grande y complejo, la comunicación de corto alcance (paracrina) no es suficiente para coordinar el comportamiento de sus células. Por ello, han evolucionado conjuntos de células especializadas en la señalización entre partes del cuerpo muy separadas entre sí. Las más sofisticadas de ellas son las células nerviosas, o neuronas, las cuales típicamente emiten largas prolongaciones (axones), que entran en contacto con células diana alejadas. Cuando es activada por señales del ambiente o de otras células nerviosas, la neurona envía impulsos eléctricos (potenciales de acción) a lo largo de su axón; cuando uno de estos impulsos llega al terminal nervioso, en el extremo del axón, estimula al terminal para que segregue una señal química denominada neurotransmisor. El terminal nervioso entra en contacto con su célula diana a través de uniones celulares especiales denominadas sinapsis químicas, las cuales parecen estar diseñadas para asegurar que el neurotransmisor sea liberado sobre la membrana postsináptica de la célula diana, rápida y específicamente. Este proceso se conoce con el nombre de señalización sináptica.
*Las células endocrinas y las células nerviosas están especializadas en diferentes tipos de señalización química.
En la figura 4, se contrastan los diferentes sistemas a través de los cuales las células endocrinas y las células nerviosas coordinan el comportamiento celular en los animales.
Figura 4. Diferencia entre la señalización endocrina (A) y la sináptica (B). Las células endocrinas y las células nerviosas actúan conjuntamente coordinando las diversas actividades de los miles de millones de células de un animal superior. Las células endocrinas segregan a la circulación muchos tipos diferentes de hormonas, para señalizar células diana específicas. Las células diana tienen receptores que se unen específicamente a determinadas hormonas, de forma que tienen que "atrapar" del líquido extracelular las hormonas adecuadas. En la señalización sináptica, por el contrario, la especificidad reside en los contactos entre las prolongaciones nerviosas y las células nerviosas determinadas que señalizan: habitualmente, sólo la célula diana que se halla en contacto sináptico con la célula nerviosa está expuesta al neurotransmisor, liberado por el terminal nervioso; (a pesar de que algunos neurotransmisores actúan de una manera paracrina como mediadores locales que influyen sobre muchas células diana en una cierta área). Mientras que diferentes células endocrinas han de utilizar diferentes hormonas para conseguir comunicarse específicamente con sus células diana, muchas células nerviosas pueden utilizar el mismo neurotransmisor y, a pesar de ello, comunicarse también de una forma específica.
Dado que la señalización endocrina depende de la difusión y del flujo sanguíneo, es relativamente lenta: normalmente tras la secreción, las hormonas tardan del orden de minutos o incluso de horas en alcanzar la célula diana. Además, la especificidad de la señalización del sistema endocrino depende completamente de la naturaleza química de las moléculas señal y de los receptores de las células diana.
Las células nerviosas, por el contrario, pueden alcanzar una velocidad y una precisión mucho más elevadas. Pueden transmitir información a grandes distancias mediante impulsos eléctricos, que transportan la señal a lo largo de las prolongaciones nerviosas, a velocidades de hasta 100 metros por segundo. El impulso eléctrico se transforma en señal química local, únicamente, cuando se libera un neurotransmisor en las terminaciones nerviosas.
Las señales químicas liberadas por las células nerviosas pueden actuar de una forma paracrina o sináptica. Según el sistema paracrino, el neurotransmisor actúa como un mediador químico local, difundiendo hacia el exterior y afectando todas las células del entorno inmediato que tengan receptores para la molécula liberada. La señalización, según el sistema sináptico, es mucho más precisa; ya que los efectos del neurotransmisor quedan confinados a una sola célula diana, aunque las células adyacentes tengan receptores para este mismo neurotransmisor. En este caso, el neurotransmisor difunde no más de 100 nm de la célula diana, un proceso que dura menos de un milisegundo.
Otra diferencia entre la transmisión endocrina y la sináptica es que, mientras que las hormonas se diluyen enormemente en la sangre circulante y en el líquido intersticial, por lo que han de poder actuar a concentraciones muy bajas, los neurotransmisores se diluyen mucho menos, pudiendo llegar a alcanzar concentraciones locales altas. Por lo tanto, en la señalización sináptica, los receptores de los neurotransmisores tienen una afinidad relativamente baja para sus ligandos y, como resultado de ello, no responden de forma significativa a las bajas concentraciones de neurotransmisor que llegan a ellos desde las sinapsis vecinas.
(Además, los neurotransmisores son rápidamente retirados de la hendidura sináptica, bombeándolos de nuevo hacia el terminal nervioso o hacia células vecinas. Esta rápida eliminación asegura, no sólo la precisión espacial de la comunicación, sino también su precisión temporal. Un breve pulso de neurotransmisor liberado evoca rápidamente una breve respuesta, de forma que el patrón temporal de la señal puede ser transmitido de célula a célula).
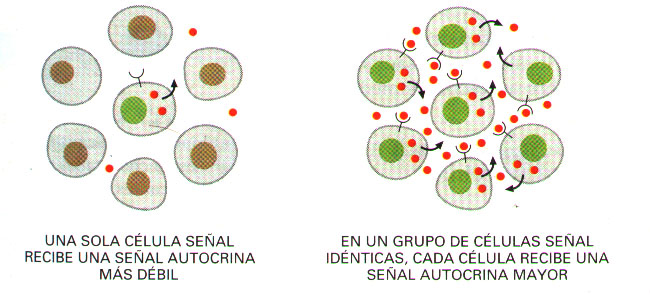
*La señalización autocrina puede coordinar decisiones de grupos de células idénticas.
Todas las formas de comunicación que hemos descrito hasta ahora, permiten a un tipo celular influir sobre otro tipo celular. Sin embargo, mediante el mismo mecanismo, las células pueden enviar señales a otras células del mismo tipo, de lo que se deduce que pueden enviarse incluso señales a ellas mismas. En una señalización de este tipo, denominada autocrina, una célula segrega moléculas señal que pueden unirse a receptores de la propia célula.
La señalización autocrina es más efectiva cuando se lleva a cabo simultáneamente por varias células vecinas del mismo tipo, por lo que puede utilizarse para estimular a grupos de células idénticas para que tomen las mismas decisiones (Figura 5).
Figura 5. Señalización autocrina. Un grupo de células idénticas produce concentraciones de moléculas señal más elevadas que una célula sola.
*Cada célula está programada para responder a combinaciones específicas de moléculas señal.
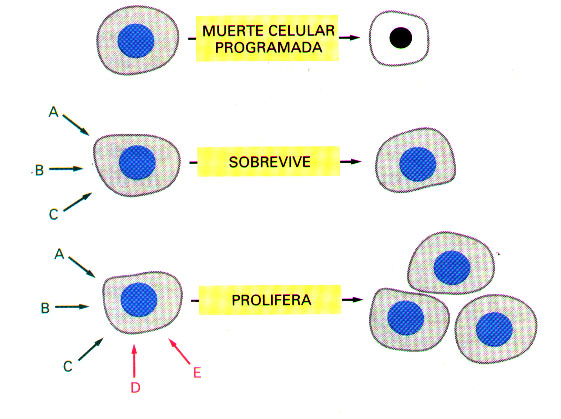
Cualquier célula dada de un organismo pluricelular está expuesta a muchas señales diferentes de su entorno. Éstas pueden actuar en millones de combinaciones posibles. La célula responderá a esta selectividad de babel, de acuerdo con su carácter específico adquirido mediante la progresiva especialización celular, en el curso del desarrollo. Así pues, una célula puede estar programada para responder a un conjunto de señales, diferenciándose; a otro conjunto de señales, proliferando; y a un tercer grupo de señales, desarrollando algunas funciones especializadas.
La mayoría de las células de los animales superiores, sin embargo, están programadas para depender de un grupo específico de señales, simplemente para sobrevivir. Cuando son deprivadas de las señales adecuadas, las células inician un programa suicida y se autodestruyen, proceso denominado muerte celular programada.
Diferentes tipos de células requieren diferentes conjuntos de señales de supervivencia, y por ello su localización está restringida a diferentes ambientes en el cuerpo (Figura 6).
Figura 6. Señalización combinatoria. Cada tipo de célula presenta un conjunto de receptores, que le permite responder a un conjunto correspondiente de moléculas señal producidas por otras células. Varias de estas moléculas señal actúan de forma combinada, regulando el comportamiento de la célula. Como se muestra en la figura, muchas células requieren múltiples señales (flechas verdes) para sobrevivir y señales adicionales (flechas rojas) para proliferar; si se depriva a las células de todas esas señales, inician un programa de muerte celular.
*Diferentes células pueden responder de forma diferente a la misma señal química.
La manera específica en que una célula reacciona con su entorno, varía, en primer lugar, de acuerdo con el conjunto de proteínas receptoras que posee la célula y a través de las que detecta un conjunto particular de todas las señales que le son asequibles y, en segundo lugar, de acuerdo con la maquinaria intracelular a través de la cual la célula integra e interpreta la información que recibe. Así, a menudo, una misma molécula señal tiene efectos diferentes sobre células diana diferentes.
No siempre las diferencias en la respuesta se deben a diferencias en los receptores. En muchos casos, la misma molécula señal se une a receptores idénticos y produce respuestas muy diferentes en distintos tipos de células diana., lo cual refleja diferencias en la maquinaria enzimática a la que están acoplados los receptores.
5. - ADAPTACIÓN DE LAS CÉLULAS DIANA.
Normalmente, cuando las células y los organismos responden a algún estímulo, pueden detectar el mismo porcentaje de variación de la señal en una gama muy amplia de intensidades del estímulo. A nivel celular, esto requiere que las células diana sufran un proceso de adaptación o desensibilización, mediante el cual su respuesta va disminuyendo cuando se halla expuesta a un estímulo durante un período prolongado de tiempo. De esta forma, la célula ajusta de forma reversible su sensibilidad al nivel del estímulo.
En el caso de la señalización química, la desensibilización permite a la célula responder a cambios de la concentración de la molécula ligando (en lugar de responder a concentraciones absolutas del ligando) en un amplio margen de concentraciones absolutas.
El principio general es sencillo: la adaptación se consigue a través de una retroalimentación negativa que actúa con un cierto retraso. La retroalimentación negativa significa que una respuesta fuerte modifica la maquinaria que produce dicha respuesta, de forma que se inhibe a sí misma; pero gracias al retraso, un cambio repentino del nivel de estímulo es capaz de producir una respuesta intensa durante un período de tiempo corto, antes de que la retroalimentación negativa tenga tiempo de actuar.
La adaptación a señales químicas puede producirse de diferentes formas. En algunos casos, se produce por la disminución progresiva del número de proteínas receptoras específicas de superficie, la cual normalmente se produce en un intervalo de horas. En otros casos se produce por una rápida inactivación de estos receptores, lo cual se produce en cuestión de minutos. En otras ocasiones, es debida a cambios en las proteínas que participan en la transducción de la señal tras la activación de los receptores, los cuales se producen habitualmente a unas escalas de tiempo intermedias.
6.-ENFERMEDADES CAUSADAS POR DEFECTOS EN LA COMUNICACIÓN INTERCELULAR.
En Medicina es, a la vez, clínicamente útil e intelectualmente satisfactorio descubrir el mecanismo responsable de la aparición de una enfermedad o grupo de enfermedades de causa desconocida. Recientemente, se ha demostrado que un cierto número de enfermedades humanas, entre ellas afecciones tan diversas como el cólera, hipertiroidismo, miastenia gravis y ciertos tipos de diabetes, surgen como consecuencia de un mecanismo común: un defecto de comunicación intercelular.
El trabajo de distintos investigadores ha permitido establecer que la clave del cólera reside en una disfunción de los receptores. W. E. Van Heyningen y colaboradores, de la Universidad de Oxford y de la Facultad de Medicina de la Johns Hopkins, demostraron que la toxina producida por la bacteria del cólera se unía a receptores de células que recubren el intestino delgado. Dos grupos de la Facultad de Medicina de Harvard establecieron que la toxina de la bacteria del cólera provocaba la secreción de líquido en el intestino delgado, (la célula bombea de 20 a 30 litros de agua hacia la luz del intestino delgado). Puesto que el líquido no puede ser reabsorbido a la misma velocidad, cantidades masivas del mismo se pierden por vómitos o diarrea. Las enormes pérdidas de líquido son la causa del gran número de muertes producidas por el cólera.
Nuestro cuerpo es una comunidad de células, en la que cada una ocupa su debido sitio donde realiza las tareas asignadas en beneficio del organismo. Salvo los leucocitos, las células normales permanecen en el tejido del que forman parte. Muy distintas son las células cancerosas, verdaderos pícaros que agreden e invaden otros tejidos.
Los 30 billones de células que forman un cuerpo normal y sano, viven en un condominio complejo e interdependiente, en el que unas regulan la proliferación de otras. Las células normales sólo se reproducen cuando reciben las instrucciones adecuadas que les envían otras células vecinas. Tal colaboración permanente asegura que cada tejido mantenga el tamaño y la arquitectura adecuada a las necesidades del cuerpo.
Las células cancerosas vulneran ese esquema de comunicación. Ignoran los controles normales de proliferación y siguen sus propias instrucciones internas de reproducción, lo que puede conllevar a la muerte del organismo.
7. -CONCLUSIONES.
En un organismo pluricelular, las células necesitan comunicarse entre sí para coordinar todas sus funciones. Las células animales se comunican mediante tres sistemas:
a) Señalización indirecta o remota por moléculas segregadas.
b) Señalización directa por contacto a través de moléculas unidas a la membrana plasmática.
c) Señalización directa por contacto vía uniones gap.
Existen tres estrategias de comunicación química intercelular, que se diferencian entre sí por la distancia a la que transmiten las señales:
a) En la señalización endocrina, las hormonas son transportadas por la sangre hasta las células diana de todo el cuerpo.
b) En la señalización paracrina, los mediadores químicos locales son rápidamente captados, destruidos o inmovilizados, de forma que únicamente actúan sobre las células situadas en el entorno inmediato de la célula que los ha segregado.
c) En la señalización sináptica, las células nerviosas segregan neurotransmisores, que solamente actúan sobre la célula diana postsináptica adyacente.
En general, podemos decir que las células se comunican entre sí liberando determinadas señales, que recorren distancias variables. Los mensajes son captados por receptores que transmiten la información a estructuras intracelulares, donde la información captada desencadena una respuesta bioquímica.
Bajo mi punto de vista, al ser la célula, la unidad estructural y funcional básica de un organismo vivo, la comunicación entre éstas resulta crucial para la supervivencia de cualquier organismo pluricelular.
8. - BIBLIOGRAFÍA.
-
ALBERTS, B y colaboradores. Biología molecular de la célula. Barcelona, Omega, S.A. (3ª ed.). (1996).
-
BARRITT, G. J. Communication Within Animal Cells. Oxford, UK: Oxford Science Publications. (1992).
-
HARDIE, D. G. Biochemical Messengers: Hormones, Neurotransmitters and Growth factors. London. Chapman and Hall. (1990).
-
MORGAN, N. G. Cell Signalling. Milton Keynes, UK. Open University Press. (1989).
-
SNYDER, S. H. Base molecular de la comunicación intercelular. Investigación y Ciencia. (Diciembre, 1985).
-
RUBENSTEIN, E. Enfermedades causadas por defectos en la comunicación intercelular. Investigación y Ciencia. (Noviembre, 1996).
-
WEINBERG, R.A. Así se produce el cáncer. Investigación y Ciencia. (Septiembre, 1992).
Breve discurso sobre comunicación intercelular
Comunicación intercelular Introducción
1
18
Comunicación intercelular Principios generales de la comunicación intercelular
Comunicación intercelular Uniones de tipo gap
Comunicación intercelular Señalización química
Comunicación intercelular Adaptación de las células diana
Comunicación intercelular Enfermedades causadas por defectos en la comunicación intercelular
Comunicación intercelular Conclusiones
Comunicación intercelular Bibliografía
Descargar
| Enviado por: | Manuel Linares |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.