Biología
Botánica
Apuntes de botánica
T22- Plantas vasculares
Ya sabemos que los embriófitos (plantas con embrión) se dividen en briófitos y plantas vasculares. Las plantas vasculares; también llamadas traqueófitos y cormófitos; tienen verdaderas raíces, tallos y hojas con verdaderos tejidos; y su desarrollo está muy ligado a la conquista del medio terrestre.
Sistemática de plantas vasculares
Dentro de las plantas vasculares tenemos:
Pteridófitos: plantas vasculares sin semilla, también llamadas “inferiores”. Entre ellos están las colas de caballo, los helechos, los licopodios…
Espermatófitos: plantas vasculares con semilla, también llamadas “superiores”,
Gimnospermas: con semilla desnuda
Coníferas
Cícadas
Ginkgo
Gnetófitos
Angiospermas: con la semilla protegida por el fruto
Monocotiledóneas (gramíneas, bambú, orquídeas).
Dicotiledóneas (rosal).
Tendencias evolutivas en el ciclo de plantas vasculares
Reducción del gametófito y pérdida de independencia del protalo femenino.
En pteridófitos, son independientes. El gametófito se llama generalmente protalo, es pequeño y de aspecto taloso. En espermatófitos la generación dominante es el esporófito, y el gametófito ha sido reducido al grano de polen y al saco embrionario.
Diferencia en dos líneas en esporangios, esporas y gametófitos
Los pteridófitos tienen isosporia: hay una única línea de esporangios, esporas y gametófitos. Los protalos son hermafroditas, el mismo da lugar a los gametos femeninos y masculinos.
Sin embargo, la tendencia en cormófitos es la heterosporia: se diferencian dos líneas de esporangios (mega y microsporangios), de esporas (mega y micrósporas) y de gametófitos (megagametófito y microgametófito). Los protalos son unisexuales, cada individuo da lugar a un tipo de gametos y el desarrollo del embrión se da en el interior del megagametófito.
Retención de gametófito en la espora y aumento de la protección del embrión
En la isosporia, el protalo se forma fuera de la espora (desarrollo exospórico) y su única protección es el tejido del protalo. En la heterosporia, el protalo se desarrolla en el interior de la espora (desarrollo endospórico) y como protección, aparte del tejido del protalo femenino, tiene la pared de la megáspora.
Hay una única excepción; un género de helechos que son heterospóricos pero tienen desarrollo exospórico.
En gimnospermas, la protección es mayor: tiene además el megasporangio y la cubierta del esporófito (testa seminal). En angiospermas tiene protección máxima: tejido del protalo femenino, pared de la megáspora, megasporangio, cubierta del esporófito y cubierta del fruto.
Pérdida de gametos flagelados (paso de oogamia a sifonogamia)
Hasta ahora y también en pteridófitos, el gameto masculino es flagelado y nada hasta el femenino. En gimnospermas aparece la sifonogamia, en la que se genera un tubo polínico que penetra las capas de las ovocélulas y llega hasta las oosferas. Así se logra una independencia del agua en la reproducción.
T23- Plantas vasculares II. Tejidos vegetales.
Características generales
Tienen el esporófito con máxima diferenciación histológica: tienen tres tipos de órganos; raíz, tallo y hojas (dos anteriores, vástago); y tres tipos de sistemas; dérmico, parenquimático y vascular. Toman medidas para la adaptación al medio terrestre:
Balance hídrico
Desarrollo de un sistema conductor eficaz tanto en absorción como en transporte; y protección para evitar pérdidas (homeohidra): epidermis cutinizadas con estomas, tricomas y tejidos aislantes secundarios como el corcho.
Calor y variaciones de temperatura
Se mejora la transpiración para compensar las oscilaciones térmicas bombeando agua fuera de forma más rápida y eficaz.
Dificultades de estabilidad por aire y viento
Como ahora no pueden flotar, se desarrollan tejidos de sostén (colénquima, esclerénquima, súber, xilema secundario) y se aumenta la superficie fotosintética para reducir la resistencia al viento (hojas tienen gran superficie, poco peso y son elásticas).
Reproducción: protección del embrión
Se retiene el gametófito femenino en la megáspora, se protege el gametófito con la pared de la megáspora, el megasporangio, la cubierta seminal y las cubiertas del gruto. Con ello aumenta la protección del embrión al que el gametófito dará lugar.
Reproducción: independencia del agua
Se pasa de oogamia a sifonogamia.
Las principales novedades de los cormófitos son:
Aumento de la diferenciación celular, aparición de tejidos.
Sistema vascular
Sistema dérmico con cutina (cutícula) y suberina (corcho)
Tejidos de sostén (lignina)
Órganos reproductores y embrión protegidos
Fecundación independiente del agua.
Tejidos vegetales. Meristemas.
También llamados tejidos embrionarios, mantienen su capacidad de diferenciación toda la vida. Los hay primarios y secundarios.
Meristemos primarios
Sus células son descendientes directas de las células embrionarias; son siempre totipotentes, y dejan poco espacio intercelular. Entre sus funciones están el crecimiento en longitud, el aumento de la superficie fotosintética, originar tejidos primarios y la reembrionalización. Aparecen en la primera temporada de la planta.
Los más importantes son los meristemos apicales, que tienen varias capas que darán lugar a diferentes tejidos:
Protodermis → epidermis
Procámbium → tejidos vasculares primarios
Meristemo fundamental → tejidos fundamentales
Otros meristemos son los remanentes, que no son apicales y que permanecen en zonas adultas para, por ejemplo, elongar o ensanchar (ejemplo importante: intercalares)
Meristemos secundarios
Sólo en dicotiledóneas y gimnospermas, y sólo en la raíz o el tallo. Se diferencian normalmente a partir de tejidos ya diferenciados. Generalmente son laterales; su función principal es la reembrionalización: células poco diferenciadas pueden volver a embrionarias. También se encargan del crecimiento en espesor.
Hay dos tipos: cámbium vascular y cámbium suberoso o felógeno.
Cámbium vascular
Da los tejidos vasculares y de sostén secundarios; se sitúa en el centro de la raíz y tallo como un anillo de células meristemáticas o en grupos aislados cuyo conjunto define un anillo. Tiene dos tipos de células:
-
Iniciales fusiformes: alargadas, dan elementos conductores de xilema y floema.
-
Iniciales radiales: más isométricas, producen las células acompañantes o elementos no conductores (parénquima radiomedular: parénquima de zonas radiales, el longitudinal es perpendicular).
-
Cámbium suberógeno o felógeno
-
Tejidos vegetales. Tejidos adultos
-
Tejido fundamental o parénquima
-
Tejidos de sostén
-
Colénquima
-
Esclerénquima
-
Fibras de esclerénquima: alargadas, aparecen en tallos y grandes hojas de monocotiledóneas. Importantes en industria (lino, etc).
-
Células de esclerénquima, esclereidas o células pétreas: muy lignificadas, en tallos y hojas. Función protectora y de sostén, presentes en la corteza de plantas leñosas y frutos.
-
Tejido asilante: epidérmico
-
Tejidos conductores o vasculares
-
Xilema o leño
-
Floema o líber
-
Células conductoras: anucleadas.
-
Células cribosas
-
Tubos cribosos
-
Células no conductoras
-
Células acompañantes (albuminíferas o anexas)
-
Refuerzos (parénquima, esclerénquima)
-
Haces vasculares
-
Concéntricos: normalmente el xilema está dentro (anficribal); en otros, está por fuera (anfibasal).
-
Colaterales: no concéntricos. Puede haber distintos tipos según la disposición de xilema, floema y cámbium.
-
Estela
-
Protostela
-
Sifonostela
-
Eustela
-
Crecimiento secundario
-
Tejidos glandulares
-
Protección: mediante secreciones repelentes o venenosas para hongos o herbívoros; o secreciones que cierran heridas y evitan infecciones.
-
Atracción de polinizadores por aceites esenciales o recompensas: glándulas nectaríferas.
-
Florales: polinización
-
Extraflorales: defensa
-
Atracción de insectos (plantas carnívoras)
-
Conductos laticíferos
-
Conductos resiníferos
-
Cavidades oleíferas
-
Estructura y desarrollo del cormo
-
Origen del tallo: la semilla
-
Funciones del tallo
-
Conducción
-
Soporte (de otros tallos, hojas, flores…)
-
Almacenamiento (rizomas, tubérculos)
-
Fotosíntesis en tallos jóvenes y de herbáceas (cáctus, etc).
-
Crecimiento del tallo
-
Tipos de yemas
-
Según posición: apicales, laterales, adventicias
-
Apicales: terminales.
-
Laterales: pueden ser axilares, situadas en los nudos; o supletorias/accesorias, situadas junto a las axilares. Dentro de ellas distinguimos las colaterales (formando una fila en horizontal) y las seriales (fila vertical).
-
Adventicias: en raíz, hoja o entrenudo. Sin relación con meristemos apicales.
-
Según estructura: desnudas, peruladas
-
Según comportamiento: durmientes, activas
-
Según función o significación: floríferas, foliíferas o mixtas
-
Según situación: haplo, diplo y triplocaulescente.
-
Ramificación del tallo
-
Tipo de ramificación
-
Dicotómica: el ápice se divide en dos por división de la célula apical, y se da lugar a dos ramas. En otros casos el ápice cesa de crecer, en la periferia se diferencian 2 células en células apicales y cada una organiza un nuevo ápice (mismo resultado). Se da en algunas pteridófitas.
-
Lateral: Las ramas se originan en yemas axilares, a partir de la segunda o tercera hoja desde el ápice. Predominante en espermatófitas.
-
Monopodial:

típica de coníferas. Todo el sistema es atravesado por un eje principal único o monopodio, con crecimiento indefinido. El eje principal crece más intensamente que los ejes laterales de primer orden, y estos que los de segundo orden, y así sucesivamente. -
Simpódica: las ramas laterales se desarrollan más que el eje principal. El eje madre puede incluso interrumpir por completo su crecimiento porque su yema apical quede en reposo, se transforme en una flor o muera. Entonces las yemas axilares (generalmente las superiores) continúan el
-
Dicasio: cuando son dos las ramas laterales del mismo orden que continúan el crecimiento (suelen estar opuestas entre sí).
-
Monocasio: cuando la ramificación se continúa constantemente por una sola rama lateral. Con frecuencia esta rama se dispone en la prolongación de su brote madre, desviando el extremo de éste hacia un lado. Así se origina un sistema de ramificación con un eje aparente, pero que se compone de ramas laterales de orden diferente.
-
Filotaxis
-
Grado de ramificación
-
Potencialidad de las ramas
-
Acrotonía: las ramas superiores favorecidas frente a las inferiores.
-
Basitonía: las ramas inferiores favorecidas. Siempre en arbustos.
-
Mesitonía: las ramas del medio favorecidas.
-
Epitonía: favorecidas las ramas del lado superior
-
Hipotonía: favorecidas las del lado inferior.
-
Anfitonía: favorecidas las que dan ramas horizontales.
-
Braquiblastos: con entrenudos cortos y crecimiento definido
-
Macroblastos: con entrenudos largos y crecimiento indefinido.
-
Desarrollo ontogenético del tallo
-
Estructura interna primaria del tallo
-
Estructura interna secundaria del tallo
-
Crecimiento en longitud. Xilema y floema secundarios
-
Crecimiento en grosor. Peridermis
-
Súber: células muertas sin espacios intracelulares. Las paredes están recubiertas de suberina, sustancia que lo hace impenetrable al agua y los gases. Las lenticelas son células suberosas más o menos sueltas que permiten el intercambio de gases.
-
Felodermis: a diferencia del cámbium que genera una capa tras otra, cesa su actividad cada año, formándose una nueva felodermis al año bajo la del anterior. La antigua queda aislada del resto como tejido muerto. El conjunto de capas de peridermis es el ritidoma o la corteza externa.
-
Partes de la madera
-
Anillos de crecimiento
-
Crecimiento secundario en monocotiledóneas
-
Filomas
-
Cotiledones: los primeros órganos homólogos a aloja que salen. Son hojas embrionarias, ¿fotosíntesis? En monocotiledóneas hay un solo cotiledón, en dicotiledóneas dos, en gimnospermas muchos.
-
Catáfilos: de aspecto membranoso o coriáceo, hay varios tipos (hojas de la cebolla). Con misión protectora, son el único tipo de hojas de los rizomas.
-
Nomófilos: hojas en sentido estricto. Verdes, intervienen en la fotosíntesis y en la respiración. Hay muchos tipos, y sirven para clasificación taxonómica.
-
Hipsófilos o brácteas: hojas estériles que sirven para protección o atracción (corolas, hojas de buganvillas).
-
Antófilos: hojas de la flor (modificadas): sépalos, pétalos, estambres y carpelos.
-
Crecimiento de la hoja
-
Morfología de la hoja
-
Limbo o lámina: parte más importante, bifacial; formada por haz (cara adaxial, la más cercana al eje) y envés (cara abacial, la más lejana del eje). Sus extremos se denominan base del limbo y ápice, lo recorre el nervio principal o medio y puede tener también nervios secundarios.
-
Peciolo: rabito que une el limbo con el tallo
-
Base foliar: el punto de inserción donde se une con el tallo, encima está la yema axilar. Puede ser simplemente un ensanche o estar formada por estípulas.
-
Nerviación
-
Palmatinervias: de la base del limbo salen varios nervios centrales.
-
Paralelinervias: no hay un nervio central, sino varios paralelos de igual importancia. Común en monocotiledóneas.
-
Pinnatinervias: nerviación como una pluma; la más común en dicotiledóneas.
-
Contorno del limbo
-
Margen
-
Hojas simples
-
Enteras: borde liso
-
Divididas: dentadas (con pequeños dientes en el margen), aserradas (todos los dientes miran al ápice de la hoja), crenadas (dientes más pequeños), lobuladas (mucho más grandes):
-
Hendidas (-tífidas): eje no llega a la mitad
-
Partidas (-partidas): eje llega a más de la mitad
-
Seccionadas (-sectas): eje llega al raquis pero no se hace independiente.
-
Hojas compuestas: con varios foliolos de limbo que quedan adheridos al raquis (parecido al tallo, pero diferente porque no tiene yemas)
-
Pinnado-compuestas: con un eje central
-
Paripinnadas: número de foliolos pares
-
Imparipinnadas: foliolos impares, con uno apical.
-
Palmeado-compuestas: foliolos anexos al comienzo del peciolo
-
Trifoliadas: caso intermedio
-
Ápice
-
Base del limbo
-
Tipo de inserción de la hoja
-
Evolución pteridófitos
-
Características diferenciadoras de briófitos
-
Predominio del esporófito
-
Haces conductores (lignina)
-
Verdaderas raíces
-
Cutícula bien desarrollada
-
Estomas
-
Gametófito muy reducido e independiente del esporófito
-
Plantas vasculares primitivas
-
Evolución a partir de plantas vasculares primitivas
-
Ramificación lateral: por culminación o sobrecrecimiento, un eje domina sobre otro, el cual acaba siendo una ramificación lateral.
-
Hojas grandes: por aplanamiento de una rama de crecimiento corto cuyos ejes (acaban siendo nervios) se unen por un tejido.
-
Hojas pequeñas: por reducción de las ramas.
-
Eustela: por concrescencia se juntan varios telomas alrededor de un círculo.
-
Flores: por incurvación de esporangios terminales.
-
Estructura de pteridófitos. Esporófito.
-
Haces vasculares
-
Crecimiento secundario
-
Hojas
-
Origen de micrófilos
-
Tipos de hojas
-
Trofófilos: hacen la fotosíntesis, tienen importancia trófica.
-
Esporófilos: dedicadas a la reproducción, tienen esporangios y esporas.
-
Trofoesporófilos: hacen las dos cosas; parte de la hoja se dedica a los soros y parte a la fotosíntesis.
-
Esporangios y esporas
-
Estructura de pteridófitos. Gametófito (protalo)
-

Clase Psilotópsida -
Orden Psilotales
-
Orden Ophioglossales
-
Clase Equisetopsida
-
Orden Equisetales
-
Familia Calamitaceae
-
Familia Equisetaceae
-
Reproducción
-
Clase Marattiopsida
-
Orden Marattiales
-
Familia Marattiaceae
-
Clase Filicopsida
-
Tallo y raíz
-
Hojas
-
Esporangios (leptosporangios)
-
Desnudos: sin indusio
-
Con indusio lateral
-
Con indusio central
-
Con pseudoindusio: hoja enrollada.
-
Orden Salviniales
-
Familia Marisiliales
-
Familia Salviniaceae
-
Orden Filicales?
-
Distribución y modo de vida de Pteridófitos
El otro tipo es el cámbium suberógeno o felógeno, que forma tejidos protectores: epidermis o corteza secundaria (súber o corcho hacia fuera; felodermis hacia dentro, puede no estar). Se forma en posición externa a la planta, inmediatamente debajo o muy cerca de la dermis, y tiene duración anual.
No se dividen ni crecen.
Presente en todos los órganos de la planta, en los dos tipos de crecimiento. Es un tejido primario de células grandes y alargadas con paredes finas y espacios intercelulares. Están poco diferenciadas, podrían reembrionalizarse.
Aparece en el relleno del córtex (la parte de parénquima más alejada de la médula, al otro lado de los haces vasculares) y en la médula de tallo y raíz, en el mesófilo de la hoja, en la pulpa de los frutos, en las semillas (en todas partes). Según su situación desempeña distintas funciones; interviene en la fotosíntesis (clorofílico), en la respiración (aerífero) y en la reserva de almidón, lípidos, agua o aire (de reserva).
Hechos por células de paredes engrosadas, rígidas y resistentes a la presión.
Proporciona sostén mientras crece en longitud (primario) y en herbáceas. Después desaparece o se transforma en otros tejidos. Sólo en los órganos aéreos de la planta, próximo a la epidermis. Está formado por células vivas alargadas con pared primaria muy engrosada. Rodea los vasos conductores.
Proporciona sostén en órganos que han parado de crecer. Formado por células maduras sin citoplasma (muertas); con paredes engrosadas y frecuentemente las secundarias lignificadas. Tiene dos tipos de células:
Tejido que contacta con el medio, normalmente monoestratificado. Sin espacios intercelulares y con paredes secundarias especiales: cutículas (formadas por cutina. Sólo en la superficie libre de la célula y por encima de las primarias). No hay en la raíz, y sirven para proteger ante los hongos. A veces también tienen ceras impermeabilizadoras, y a veces por encima se produce una silificación/calcificación (plantas rasposas). La cutícula no se descompone, así que queda en los fósiles.
Hay dos especializaciones importantes de la epidermis: estomas y tricomas.
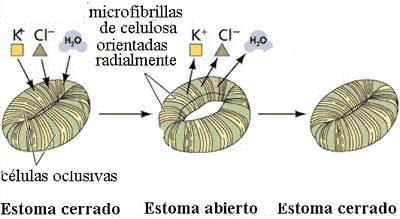
Los estomas son poros rodeados por dos células (oclusivas) que según se hinchen, se abren o cierran. Las únicas células de la epidermis con cloroplastos.
Los pelos o tricomas pueden ser uni o pluricelulares, y tienen muchas funciones: soporte de plantas trepadoras, disminución de la transpiración, defensa ante depredadores, diseminación de frutos, pelos urticantes, percepción de estímulos, secreción, absorción (pelos radicales), etc.
Si hay crecimiento secundario, la epidermis es sustituida por la peridermis; sólo en raíces y tallo de dicotiledóneas y gimnospermas. Se forma a partir del cámbium suberoso o felógeno, que se diferencia hacia el exterior en células de súber o corcho (paredes celulares con suberina, impermeable) y hacia el interior felodermis (no siempre). Como el felógeno tiene actividad anual, cada vez se origina en capas más profundas; por lo que llega a aparecer entre el floema no funcional y lo empuja (elimina)
En esta peridermis no encontramos estomas, sino lenticelas; que son también poros pero no tienen capacidad para cerrarse.
Son tejidos completos. Las células están orientadas longitudinalmente, y hay líquidos que circulan por el interior de las células.
Transporta agua y nutrientes inorgánicos hacia las hojas, es más duro que el floema. Está formado por células conductoras o elementos traqueales, (muertas y lignificadas: tráqueas y traqueidas) y células no conductoras (células del parénquima y fibras xilares).
Las tráqueas o elementos de los vasos predominan en angiospermas. Formados por células muertas dispuestas en filas; se comunican por punteaduras entre filas y por perforaciones en de la misma fila. Hay gran variedad de formas de engrosamientos de la pared: anulares, helicoidales, reticuladas, punteadas…
Las traqueidas predominan en gimnospermas y pteridófitos. Constituidas por células fusiformes muertas dispuestas en filas; sólo se comunican por punteaduras, tanto entre filas como dentro de la misma fila. Su engrosamiento (lignificación) no es uniforme.
Distribuye la savia elaborada (compuestos orgánicos) por toda la planta, y sólo es funcional un año. A diferencia del xilema, como no es resistente, no da fortaleza a la planta; por lo que el floema no funcional es eliminado. Formado por:
Las células cribosas son células largas con paredes terminales inclinadas. Tienen áreas cribosas en las paredes laterales y sobre todo en las terminales. Su metabolismo está regulado por las células albuminíferas. No están huecas, así que los elementos fotosintéticos pasan de una célula a otra mediante prolongaciones citoplasmáticas. Aparecen en pteridófitos y gimnospermas.
Los elementos de los tubos cribosos comunican las paredes terminales a través de placas cribosas, formadas por una o varias áreas cribosas (simples o compuestas). Su metabolismo está regulado por las células anexas. Aparecen en angiospermas.
Tanto áreas como placas cribosas pueden ser obstruidos por calosa, que se deposita a la llegada del otoño (por eso el floema es funcional un solo año), lo que provoca la caída de las hojas.
Hay varios tipos de haces vasculares.
También distinguimos entre cerrados (ya no van a crecer más) y abiertos (sí) según posean cámbium o no (crecimiento secundario).
Patrón que presentan los haces de xilema y floema en el corte transversal del tallo, raíz o pecíolo de las plantas. Hay tres tipos básicos: protostela, sifonostela y eustela.
También llamado haplostela. Columna sólida de tejidos vasculares ubicada en posición central. Es el tipo más simple, y el más primitivo filogenéticamente. Se encuentra en algunas Pteridófitas, en angiospermas acuáticas, y en raíces primarias de plantas con semilla. Cuando la columna de xilema tiene forma estrellada en transcorte, recibe el nombre de actinostela.
El sistema vascular tiene forma de tubo, envolviendo una médula parenquimática. Puede tener lagunas foliares o no, y puede ser ectofloemática (floema por fuera) o anfifloemática (floema dentro y fuera). Si tiene lagunas foliares muy grandes y en corte transversal el anillo es discontinuo, se trata de una dictiostela.
El sistema vascular consta de haces vasculares ordenados en anillo alrededor de una médula. La atactostela es una variante característica de monocotiledóneas, con haces vasculares colaterales o concéntricos esparcidos regularmente en todo el tallo.
Sólo en gimnospermas y dicotiledóneas. Implica un aumento de diámetro del sistema vascular al cámbium vascular y la sustitución de la epidermis por la peridermis.
Producen secreciones (productos útiles para el productor) o excreciones (desechos, sustancias perjudiciales). Las funciones de los tejidos glandulares son:
Producen látex, material de desecho que tiene función cicatrizante. También es disuasorio de agresión. Contiene carbohidratos, ácidos orgánicos, alcaloides, resinas, caucho… Los tubos pueden ser articulados (varias células) o no articulados (una).
Acumulan resina (una mezcla de terpenoides) en los espacios intercelulares (conductos esquizógenos: para formarlos las células contiguas se separan). La resina sirve como reserva y ayuda a la cicatrización, la tienen las coníferas.
Almacenes de aceites esenciales (cavidades esquizógenas). Presentes en el hipérico, el eucalipto y la piel de los cítricos. Sirven como reserva y contienen compuestos tóxicos.
T 24- Plantas vasculares III. Desarrollo del cormo.
El cormo es el conjunto formado por raíz, tallo y hojas. Estos dos últimos forman el vástago. El tallo o tronco es también llamado eje caulinar; tiene crecimiento indefinido, está más o menos ramificado y está provisto de hojas. Se diferencia de la raíz por la presencia de entrenudos en los que se insertan las yemas auxiliares y las hojas, y por su fototropismo positivo. Las hojas son ramas planas laterales de crecimiento y duración indefinidos.
La semilla está compuesta de tegumento y germen o embrión. La parte que primero crece es la radícula, situada en la parte inferior del embrión y procedente de meristema apical radical. Crece por fototropismo negativo y geotropismo positivo; y una vez fuera se convertirá en una auténtica raíz.
La plúmula es una yema situada en el extremo contrario a la radícula, deriva del meristemo apical caulinar. El hipocótilo es el espacio entre radícula y plúmula; y se extiende para que la semilla emerja del suelo. Tras ello los cotiledones (hojas que contienen sustancias de reserva) se abren y la plúmula da lugar al sistema caulinar o tallo (el situado por encima de ellos es el epicótilo, por debajo sigue siendo hipocótilo). Finalmente los cotiledones se marchitan y nuevas hojas surgen del tallo.
El crecimiento del tallo se da por el desarrollo de yemas (apicales, en el extremo; o axilares, en cada nudo). Las yemas pueden estar protegidas por unas hojitas llamadas primordios foliares, que secretan mucílagos o resinas y protegen de la desecación. El tallo crece por el ápice del vástago gracias a la yema apical; las yemas axilares dan lugar tres cosas: a ramas con hojas, a ramas con flores o a flores.
En la planta aparecen nudos, puntos donde se insertan las hojas en el tallo; y entrenudos, espacios entre nudos distintos, Los entrenudos poseen meristemos intercalares, que les permiten crecer de forma regular o irregular. En plantas en roseta, los entrenudos se reducen al mínimo. En el otro extremo, si la planta no dispone de luz (aislamiento), crece todo el rato buscándola y los entrenudos son muy largos.
Normalmente clasificamos las plantas según las hojas; pero en zonas de inviernos largos donde normalmente no haya hojas, se puede utilizar otro criterio: las yemas.
Pueden ser desnudas (desprovistas de escamas protectoras, protegidas por hojas jóvenes) o peruladas (protegidas por hojas modificadas con aspecto escamoso, generalmente oscuras y coriáceas).
Pueden ser durmientes (no dan ramas en varios años o nunca) o activas; dentro ellas hay anuales (ramas de enriquecimiento en herbáceas anuales) e hibernantes (en leñosas).
La caulifloria trata el desarrollo tardío de yemas durmientes que quedan en la corteza
Las floríferas dan flores, las foliíferas hojas, y las mixtas dan una flor y una hoja.
Según se sitúen en el eje principal o tallo (haplocaulescente, pues dan lugar a ese tipo de ramificación), en las ramas secundarias (diplo) o en las ramas de las ramas (triplo).
{kind=link}
Es una estrategia que siguen las plantas para aumentar la superficie fotosintética. La configuración del vástago depende del tipo de ramificación, de la filotaxis, del grado de ramificación y de la potencialidad de las ramas.
Estos procesos están relacionados con la dominancia apical: el efecto inhibidor de la yema apical sobre las laterales.
crecimiento y la ramificación.
Dentro de la ramificación lateral simpódica podemos distinguir dos modelos distintos:
Un dicasio puede parecer una dicotomía, pero las ramas laterales no se disponen en un plano sino en distintas direcciones del espacio. Un monocasio puede parecerse al monopodio cuando el eje aparente crece recto, y los extremos de las ramas que detienen su desarrollo se disponen de modo semejante a ramas laterales.
La filotaxis es la disposición de las hojas y las yemas laterales en el tallo. Pueden estar opuestas o alternas.
En la ramificación haplocaulescente las hojas salen directamente del eje central, no hay ramas. En la diplocaulescente salen en las ramas laterales, hay eje secundario. En la triplocaulescente salen en las ramas de las ramas; y así sucesivamente.
Algunas ramas crecen más que otras en el mismo tiempo (anisocladia: dos tipos de tallo por crecimiento). Así, distinguimos varios tipos de ramas:
Según su crecimiento y situación:
En el mismo lugar, según crecimiento, inducida por gravedad:
En el mismo lugar:
Entre los macroblastos están los escapos (vástagos largos sin ramificación ni hojas) y los estolones (tallos largos que crecen y enraizan).
En el crecimiento primario, los meristemos primarios dan lugar a los tejidos primarios: la protodermis, a la epidermis; el procámbium, floema y xilema primarios, además de cámbium fascicular; y el meristemo fundamental los tejidos fundamentales, que dan lugar a médula, radios (dan cámbium interfascicular) y corteza (da felógeno).
En el crecimiento secundario, los meristemos secundarios dan lugar a los tejidos secundarios: el cámbium vascular (procedente de cámbium fascicular e interfascicular) da lugar a floema y xilema secundarios, y el felógeno a la peridermis (formada por súber y felodermis).
Lo vemos de fuera a adentro. Hay varios tipos según la disposición del haz vascular (protostela, sifonostela…).
Primero encontramos la epidermis, con cutícula, estomas y tricomas. Luego viene el córtex, zona parenquimática y de sostén. Después, una banda llamada endodermis, que tiene bandas de Carpay que seleccionan qué entra y qué sale al cilindro vascular. Finalmente, el cilindro vascular, con xilema y floema. En algunos casos tiene una parte central llamada médula, que puede faltar (dejando canal medular).
Normalmente el xilema va por dentro del floema. El cámbium vascular, que da lugar a xilema y floema secundarios, se dispone en haces; el fascicular es el que corresponde a esos haces y el interfascicular a las zonas sin haces. Si es continuo, forma un anillo meristemático.
Además del crecimiento en grosor hay otro meristemo secundario, el cámbium suberógeno. Al crecer el tallo, se rompe la epidermis y quedan restos en la parte más externa. Al nivel del córtex comienza su actividad, dando el súber fuera y la felodermis por dentro; en conjunto dan la peridermis.
Un caso particular es el corcho. En el alcornoque, el felógeno es activo forma indefinida; no se forman capas nuevas cada año a menos que quitemos el corcho.
Hay dos partes: el duramen y la albura. El duramen es el corazón de la madera; formado por tejidos muertos (traqueadas, tráqueas y fibras), no es funcional y acumula resinas, gomas y taninos que dan colores oscuros y evita que se pudra. La albura es la parte externa, contiene células vivas y es una madera más blanda y menos resistente.
El duramen y la albura están formados por xilema, luego va el cámbium vascular; luego el floema y finalmente la corteza.
Una vez tenido el crecimiento secundario, se ven los anillos de crecimiento. Cada anillo anual tiene dos bandas, que se deben al crecimiento rápido en primavera y a otro más lento en verano (queda más concentrado). A veces queda un anillo anual múltiple, debido a cambios de condiciones. La dendrocronología estudia estos anillos.
Suelen tener sólo crecimiento primario porque son herbáceas, pero a veces se da un engrosamiento especial de los tejidos primarios: se transforman en el meristemo cambial, que al exterior da una capa de células parenquimáticas y al interior abundante parénquima secundario con haces vasculares secundarios dispuestos de manera difusa.
T24b- Desarrollo del cormo. La hoja
La hoja es el órgano lateral del vástago de origen exógeno, de crecimiento y duración limitados, generalmente laminar y con simetría dorsiventral.
Sin embargo, existe el término filoma para referirse a todos los órganos homólogos a la hoja porque surgen igual, y aparecen en la planta en una sucesión a lo largo del tiempo (sucesión foliar):
La hoja se origina a partir de yemas y sufre dos tipos de crecimiento: acroplástico (se alarga) y basiplástico (se ensancha). Normalmente el acroplástico termina dejando el basiplástico, pero en los helechos se mantiene (prefoliación circinada).
Las partes de una hoja son:
El limbo es la parte por la que se identifica a las hojas, pues es la que presenta más variaciones. Entre ellas están la nerviación, el contorno del limbo, el margen, la forma del ápice, la forma de la base del limbo y el tipo de inserción de la hoja.
La hoja puede tener distintas formas: acicular (aguja), linear (más ancha, márgenes paralelos, abeto), oblonga (largamente elíptica y de bordes paralelos), ovada (huevo), elíptica, lanceolada, espatulada, deltoide (triangular), romboidal, flabeliforme (abanico, Gingko), reniforme, sagitada (punta de lanza), panduriforme (forma de guitarra), escuamiforme (espárrago).
Distinguimos entre:
Todas pueden ser pinnatinervas o palmatinervas (palmatífidas, pinnatisectas…).
Pueden ser bipinnadas si están divididas dos veces, tripinnadas…
En muchos casos, la forma del ápice tiene una función especial.
Cuneada (cuña), redondeada, asimétrica, sagitada, hastada (alabarda), etc.
T26- Pteridófitos I
Los pteridófitos también se denominan criptógamas vasculares, pues son los primeros cormófitos pero no tienen flor. Hay dos líneas evolutivas: la de los licófitos, y la de los helechos y equisetos.
Varias características los diferencian, procedentes de la adaptación a ambientes secos:
Dominan el devónico y mueren a su final. Hay tres grupos: Rhyniophyta, Zosterophyllophyta y Trimerophyta. Tienen ejes dicotómicos, esporangios apicales o laterales, protostela (haplo o actinostela) y todas son isospóreas.
Teoría del origen del cormo: teoría telomática.
¿Cómo de un eje dicotómico se diferencian hojas, flores, etc?
Están formados por tallos, hojas y raíces. Los tallos son generalmente subterráneos (rizomas). En las hojas tienen megáfilos y micrófilos; todo con sistema vascular con traqueidas (no tráqueas aún): La raíz principal suele morirse y las que siguen son las caulógenas.
En grupos extintos había protostela (haplo o plectostela), también en Psilotum y en licopodios. En los demás hay sifonostela, con lagunas foliares (helechos) o sin ellas (Selaginella). Hay varios tipos de traqueidas, desde anulares hasta escaleriformes.
Normalmente sólo hay crecimiento primario; sólo encontramos secundario en plantas vasculares sin semilla y no es indefinido. Como las células no pueden dividirse en horizontal, llega un momento en que no se puede ensanchar más; por tanto la longitud de las células es el factor limitante.
Hay dos teorías para la formación de los micrófilos; por reducción de megáfilos o por la teoría de la enación: una pequeña protuberancia del tallo es vascularizada por una traza foliar (haz que vasculariza la hoja y la conecta con el sistema vascular central del tallo). Evolutivamente podrían darse ambas.
Por una parte, puede haber micrófilos o megáfilos. En los micrófilos no hay lagunas foliares, en los megáfilos sí.
Por otra, podemos distinguir entre:
Los esporangios pueden situarse en la axila foliar o en el envés o margen; y pueden ser de dos tipos: eusporangios (pared con varias capas de células, sentados) o leptosporangios (una capa, pedunculados).
Las esporas pueden ser isósporas (dan gametófito bisexual) o heterósporas, a saber micrósporas (gametófito masculino) o macrósporas (femenino).
Lámina verde, a veces de forma acorazonada; que se fija al sustrato mediante rizoides unicelulares. Tiene una vida autónoma del esporófito, pero de apenas unas semanas. Puede ser bisexual o heterosexual, y poseer anteridios y/o arquegonios, que darán micrósporas y macrósporas.
T28- Pteridófitos III. Helechos y equisetos
La división Polypodiophyta tiene 11.000 especies, es el segundo grupo de plantas vasculares más variado tras angiospermas (más que gimnospermas). La mayoría son epífitas de ambientes tropicales, y pueden ser desde plantas pequeñas hasta tener portes arborescentes. No tienen cámbium vascular (excepto un género). Se les separa en dos grupos según el tipo de esporangios que tengan: eusporangios o leptosporangios.
Ambos tienen un desarrollo diferente.
Los eusporangios proceden de varias células epiteliales, que se dividen para dar algunas la parte estéril del esporangio, la pared (de varias capas); y otras la parte reproductiva o fértil. En el interior se diferencian las células madre de las esporas y las células que dan lugar al tapete, tejido monoestratificado que inicialmente es nutricional y que a medida que se degrada secreta sustancias que se depositan en la superficie de la espora, dándole ornamentación. Las esporas, al proceder de meiosis, se disponen en tétradas. Tienen dehiscencia normal.
Los leptosporangios proceden de una única célula, que se divide en dos: célula interna y externa. Luego la interna se divide en célula apical e interna. La célula externa da el pedúnculo, la apical la cubierta del esporangio y la interna al tapete (diestratificado) y a las células madre de las esporas. Las esporas se disponen en tétradas.
Su dehiscencia es rara, pues gracias a una serie de refuerzos en sus paredes que se contraen al secarse, el esporangio se abre y salen las esporas.
EUSPORANGIADOS
Tienen esporófito con ramificación dicotómica y micrófilos; poseen protostelas de tipo actinostela y unos rizomas (tallos subterráneos paralelos al suelo y engrosados por sustancias de reserva) micorrizados por Zygomycota, de los que salen rizoides (estructuras pequeñas que funcionan como raíces, uni o pluricelulares y que salen en cualquier sitio). Los esporangios están agregados de tres en tres en sinangios, y dan un solo tipo de esporas (isospóreos).
El gametófito es bisexual y similar al rizoma: subterráneo con simbiosis con hongos, de aspecto ramificado y (si dura más) puede estar vascularizado. Los espermatozoides son pluriflagelados, y el embrión tiene desarrollo exoscópico.
Sólo hay una familia, Psilotaceae; con dos géneros: Psilotum y Tmesipteris.
Aunque Psilotum tiene un aspecto similar a los riniófitos, se debe a una reducción secundaria de raíz y hojas. Tienen un cilindro vascular no ramificado, y están muy relacionados con Ophiglossales. Son tropicales y subtropicales, pero en la Península tenemos una especie: Psilotum nudum.
Tmesipteris se encuentra en Australia y el Pacífico y es epífito de helechos arborescentes. Tiene haz vascular no ramificado y micrófilos; es como Psilotum pero con micrófilos más grandes.
Helechos eusporangiados de pequeño tamaño, con sifonostela o dictiostela. Tienen frondes solitarias no circinadas, separadas en dos segmentos: uno fértil y otro estéril. Pueden ser enteras o divididas. Sabemos que los Ophioglossales divergieron tempranamente de otros helechos.
Los esporangios tienen dehiscencia transversal (la mayoría longitudinal), y el gametófito es heterosexual, micorrizado, subterráneo y tuberoso (en esto se parece a Psilotum). Hay dos familas: Ophiolosssaceae y Botrichyaceae.
Las primeras contienen el género de plantas con mayor número conocido de cromosomas (O. reticulatum). Las Botrichyaceae tienen el único género de helechos con crecimiento secundario, Bothychium.
Tuvieron su máximo esplendor a finales del Paleozoico. Hay dos familias: Calamitaceae y Equisetaceae.
Helechos arbóreos que llegaron a los 18 m de altura y los 45cm de grosor. Fueron las únicas plantas del carbonífero con reproducción vegetativa (por rizomas); eran fundamentalmente hidrófilos y la mayoría isospóreos. Extintos.
También llamadas colas de caballo. Sólo hay un género, Equisetum, con 15 especies. El género extinto Equisetites era muy similar a Equisetum, por eso se dice que Equisetum es un fósil viviente.
Con origen cretácico, todas son herbáceas perennes; suelen vivir en ambientes palustres (alta humedad) de todo el mundo y están conectadas con Marattiales.
Tienen ramificación verticilada (alrededor del tallo, al mismo nivel) y sus profundos rizomas les da gran capacidad de propagación, de los nudos de estos rizomas salen las raíces secundarias. Las hojas son micrófilos, aunque se interpretan como macrófilos reducidos. Las hojas se fusionan formando una vaina en verticilo; y en el nudo sale una rama por cada hoja.
Los tallos son fotosintéticos, verdes y articulados (compuestos por nudos sólidos y entrenudos largos, frágiles). También tienen estrías, que se deben a acumulaciones de colénquima (costillas o cárinas) que dan resistencia al tallo. En las zonas que no tienen costillas, llamadas valles o valéculas, se sitúan los estomas.
Los tallos son huecos, con canales por los que circula el aire; de dos tipos: valeculares (si están en la zona de valéculas, más externos) o carinales (zona de cárinas, más internos). También está el canal medular en el centro. El haz vascular; que forma eustelas en los entrenudos y sifonostelas en los nudos; es quien da lugar a estos canales por degradación (en zona de córtex y en zona de cilindro vascular), al final queda entre tales haces.
Además, en la parte exterior se deposita sílice; que les da tacto áspero, les sirve de refuerzo y sirve de protección ante hongos e insectos.
Los esporangios se agregan en número de cinco o diez en esporangióforos peltados y agrupados en estróbilos en el ápice del tallo. Tienen dehiscencia longitudinal y dos grupos de especies: en unos, se diferencian tallos estériles de fértiles (no fotosintéticos, no ramificados); en otros está todo en uno. Si son diferentes, primero salen los fértiles. Los equisetos gastan las reservas de los rizomas en la reproducción; luego germinan los tallos fotosintéticos y se dedica el resto del año a engrosar los rizomas para el siguiente.
Sus esporas son muy características por la presencia de eláteres, que se enrollan sobre ella al estar mojados pero se despliegan al secarse (eláteres higroscópicos, reaccionan al agua). Los eláteres de esporas distintas se entrelazan y se dispersan en grupos. Tienen capacidad de germinación limitada (5-7 días), lo que disminuye la viabilidad de las esporas y las hace capaces de germinar sólo en sitios muy húmedos.
El gametófito es verde, pequeño y linear. Tiene reproducción sexual limitada a zonas recientemente inundadas y ricas en nutrientes; y puede ser de dos tipos: masculino o bisexual (madura primero el arquegonio). Tienen embrión exoscópico y espermatozoides multiflagelados.
Incluye Psaronius, que fue dominante en el Carbonífero. Tienen eusporangios aislados o en sinangios (estructuras donde se fusionan varios), viven en el trópico húmedo y tienen grandes frondes pinnadas. Poseen protalos de larga vida, y los géneros más importantes son Angiopteris y Marattia.
LEPTOSPORANGIADOS
Son los helechos típicos. Unas 10.500 especies, es la clase que da diversidad al grupo Polypodiophyta. ¾ de las especies viven en los trópicos, y tienen origen devónico. Poseen raíz, tallo y hojas.
La mayoría son rizomatosos, subterráneos y sifonostélicos. También los hay columnares en los arborescentes. En general están poco o nada ramificados, no tienen crecimiento secundario y suelen tener escamas o pelos epidérmicos, llamados páleas. Del rizoma salen raíces adventicias, la embrionaria o primaria desaparece. El rizoma puede ser largo (salen hojas separadas) o corto (salen hojas concentradas).
En los helechos arbóreos, se llegan a los 20m de altura y 45cm de diámetro. Su solidez se debe al esclerénquima que acompaña los haces vasculares, y que les hace capaz de soportar frondes de 7m, grandes y divididas.
En general, como los helechos viven en umbrías, sus hojas tienen un alto porcentaje de superficie respecto al volumen para poder captar luz. Son compuestas, divididas en pinnas; y tienen prefoliación circinada (crece más rápido la cara abaxial para proteger el ápice embrional). Pueden ser perennes o caducas según el clima. Son tóxicas, pero hay gente que come brotes jóvenes.
Las frondes se pueden clasificar en enteras (lo menos común) o divididas (uni, bi, tripinnadas…). En realidad podemos distinguir entre pinnatisectas y realmente divididas. Las frondes están formadas por peciolo y lámina, que puede estar dividida en pinnas. En este caso, desde que empieza el primer foliolo y hasta el final, el nervio medio se llama ráquis. Cada pinna se une a él con un peciólulo, si no son sentadas. Cuando la pinna se divide, sus subdivisiones se llaman pínnulas; y están unidas al ráquis primario mediante el ráquis secundario. Si hay muchas divisiones, se guarda el término pínnula para el último nivel de división de la hoja.
Hay tres tipos de frondes: trofófilos (hojas estériles), esporófilos (hojas fértiles) y trofoesporófilos (lo más común, los dos juntos).
Son isospóricos, excepto en Salviniales. Los leptosporangios normalmente se presentan en estructuras conjuntas llamadas soros, que pueden estar al aire o recubiertos por un indusio (capa incompleta protectora). También aparecen a veces unas estructuras filiformes llamadas parálisis que pueden ayudar en la protección y dispersión de las esporas. Hay distintos tipos de soros según el indusio:
Los soros pueden situarse en tres posiciones distintas. En los grupos más avanzados, aparecen en la cara abaxial de la hoja; es lo más frecuente. Cuando hay pseudoindusio, se dice que aparecen en posición submarginal. Otras veces están en posición marginal, rodeados por una estructura globosa.
Los leptosporangios tienen anillo de dehiscencia que presentan unos refuerzos internos que permiten su apertura. Cada célula del anillo tiene refuerzos en la parte interior y entre las células, pero no con el esporangio así que puede abrirse donde no tiene refuerzo. Las células por donde se rompe, el labio, son más débiles. Los helechos más primitivos tienen el anillo poco desarrollado; pero la mayoría no.
Los protalos son epígeos (no subterráneos), de aspecto foliáceo, autótrofos y algunos con forma de corazón. Normalmente efímeros, pero a veces más persistentes; en él se encuentran anteridios y arquegonios. Los espermatozoides son pluriflagelados, y el desarrollo embrionario lateral (no endoscópico ni exoscópico: la primera división es lateral, no hacia arriba ni hacia abajo). A partir de cada célula sale una parte del helecho.
Helechos acuáticos, heterospóricos y de desarrollo endoscópico, además se desarrolla dentro del esporangio (cubierta extra). Los soros se acumulan en esporocarpos o sorocarpos, que tienen en su interior los micro y macrosporangios. Las micrósporas se van y las macrósporas quedan en el esporangio.
50-70 especies, hojas flotantes en forma de trébol de cuatro hojas, con rizomas subterráneos con cámaras aeríferas (por degradación de la zona externa del córtex). Sus esporocarpos se originan a partir de la hoja, que crece hasta cubrir y fusionarse alrededor de los esporangios.
Todos flotantes, con esporocarpos originados a partir del indusio. Tienen en cada nudo dos hojas y una tercera transformada en raíz, donde aparecen los esporangios.
Se distribuyen por todo el planeta. Prefieren ambientes húmedos tropicales, pero hay especies adaptadas a condiciones de xericidad. La mayoría son homeohídricos aunque hay algunas especies poiquilohídricas (reviviscentes). Sin embargo, evitan claramente los ambientes salinos (menos Achrostichum aureum en manglares).
En casi todos los medios, los helechos entran en competencia con espermatófitos por su mismo tipo de vida: los arbóreos con palmáceas y dicotiledóneas, los equisetos con juncáceas y ciperáceas, y Salvinia con las lemnáceas. No forman comunidades excepto excepciones: Pteridium aquilinum en los márgenes de los bosques.
Descargar
| Enviado por: | Rommel |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.