Biología
Análisis mendeliano
Tema 4: Extensiones al análisis mendeliano.
Variaciones de la dominancia.
Dominancia total ! dominancia parcial y semidominancia.
Los rasgos que el analizo, uno era dominante, y tenia dominancia total. Lo que significa que tanto el homocigoto dominante como el heterocigoto manifestaban la característica. Para este tipo de dominancia es necesaria la presencia de un solo alelo dominante para que el rasgo se manifieste.
Aun así hay alteraciones. El investigador Correns en 1909, realizo sus estudios con Mirabilis Jalapa, donde se fija, él se fijaba en el color de las flores. Siguiendo la metodología mendeliana cogió dos líneas puras la roja y la blanca. Al cruzarlas observó que todos los individuos de la primera generación eran rosas y cuando los cruzo entre sí, le apareció una relación de 1:2:1 de rojos, rosas y blancos respectivamente.
En este caso había una correspondencia entre el las proporciones fenotípicas y las proporciones genotípicas. Esto sucede porque no hay una dominancia total. El heterocigoto muestra un fenotipo que es el intermedio entre los fenotipos del homocigoto dominante y el recesivo. A este tipo de dominancia se le denomina dominancia parcial o semidominancia, y hay quien le llama herencia intermedia.
También existe entre los alelos de un gen (dos, uno en cada cromosoma homologo) otro tipo de relación de dominancia. Las que denominamos dominancia y que implica que el heterocigoto se manifiestan ambos caracteres. El carácter determinado por los dos alelos del heterocigoto se refiere concretamente al fenotipo de los grupos A, B y 0.
Esta determinado por tres alelos.
0! ii.
A! IA ,IA o IA i
B! IB,IB o IB i.
AB! IA IB.
Cuando se hace siguiendo la metodología de Mendel se cruzan líneas puras por dos caracteres, los Dihibridos o dobles heterocigotos. Como consecuencia se pueden producir alteraciones en la relaciones fenotípicas dihibridas. En vez de 9:3:3:1 pueden aparecer otro tipo de combinaciones.
Mendel “Los caracteres están determinados por factores y cada carácter esta determinado por dos factores”. Hoy diríamos “cada fenotipo tiene dos alelos para un gen”
Normalmente el nombre de los genes vienen de una característica que no es normal. Al alelo habitual se le denomina alelo salvaje, que suele ser dominante pero no tiene porque serlo.
-
W+ ! alelo salvaje (ojos rojos)
-
W ! alelo mutado (ojos blancos).
En 1913 encontró una mosca que no tenia ni rojo, ni blanco, le apareció un color de ojos “eosina”, estaba producido por otra forma alelica del gen W, le llamo WE.
Era la primera vez que un gen aparecía en mas de dos formas alelicas.
En 1913 un discípulo de Morgan (Sturtevant), encontró que las relaciones de dominancia entre los alelos eran la siguientes:
-
W+> WE> W: eso quiere decir que el alelo salvaje es dominante total sobre el alelo para el color eosina y para el color blanco. Además el color eosina es dominante para el del color blanco.
A los distintos alelos de un gen se le denomina serie alelica, los alelos entre ellos pueden tener de las tres clases de dominancia que hemos descrito.
-
IA = IB > i. Esto es que IA e IB son codominantes y con dominancia total sobre i.
La importancia de la existencia del alelismo múltiple desde el punto de vista evolutivo.
Cuando solo hay dos alelos para un gen, si hay dominancia total solo hay dos posibles fenotipos. Si hay semidominancia o codominancia solo hay tres posibles fenotipos.
Por lo tanto con dos alelos para un gen solo tenemos como máximo tres posibles fenotipos. Pero a medida que aumentamos el numero de alelos aumenta, también aumenta el numero de fenotipos posibles.
Cuando hay más alelos hay más fenotipos con lo cual aumenta la variabilidad y mejor será la respuesta adaptativa.
Incompatibilidad de alelos en plantas: hay algunas plantas que se autofecundan en la naturaleza, y las que no se autofecundan, es el viento o algún insecto quien les ayuda. Para rechazar aquello que no es compatible, tienen un sistema “el alelismo múltiple”: carácter de incompatibilidad en plantas.
-
Si el alelo del grano de polen esta presente en la parte femenina de la planta hay incompatibilidad.
-
Si el alelo del grano de polen no esta presente en la parte femenina entonces el polen generara el tubo polínico y se produce la fertilización.
-
Incompatibilidad de alelos en humanos: el alelismo múltiple genera también un sistema de incompatibilidad en humanos distinto del anteriormente visto. En este caso la incompatibilidad se genera cuando no hay igualdad en los alelos. De este sistema se encarga un complejo alelico y genético que se llama complejo principal de histocompatibilidad (MHC) esto aparece en mamíferos en general y en humanos concretamente el HLA que esta en el cromosoma 6, esta generado por varios genes diferentes. Algún gen puede tener hasta 40 y tantos alelos distintos. Se genera para la protección de sustancias del exterior y que no pertenecen al organismo.
-
Se cogen moscas líneas puras de mutantes observados en USA y se cruza con moscas líneas puras observadas en Canadá, todos los descendientes presentan el tamaño normal de alas conclusión “hay complementación”.
-
Al cruzar líneas puras de mutantes de USA y de Canadá. Los descendientes tenían un tamaño anormal (muy pequeño) de alas. Conclusión “no hay complementación”.
-
El amarillo es heterocigoto.
-
El agutí es recesivo con respecto al amarillo.
-
Faltan ¼ de homocigotos dominantes.
-
Puede afectar a cualquier sexo, pero es mas frecuente en mujeres que en hombres.
-
Las mujeres cuando se ven afectadas normalmente lo hacen con menos intensidad y además muestran una mayor variabilidad en cuanto a la intensidad de la afección.
-
Un hijo de una mujer afectada independientemente de su sexo tiene un 50% de posibilidades de ser afectado.
-
Si un hombre esta afectado todas sus hijas pero ninguno de sus hijos se vera afectado.
-
Afecta principalmente al sexo masculino.
-
Los hombres afectados normalmente nacen de padres no afectados y es normalmente la madre la que transmite el gen y ella es una portadora asintomática.
-
Las mujeres pueden ser afectadas, si su padre es afectado y la madre es portadora. También puede suceder si su padre es afectado y su madre no es portadora pero tiene inactivado el cromosoma X que contiene el gen normal.
-
No hay transmisión de hombre a hombre, pero el matrimonio de un hombre afectado y una mujer portadora pueda dar la apariencia de que la transmite el hombre.
-
Afecta solo a los varones.
-
Los hombres afectados siempre tienen un padre afectado salvo que hay una mutación de novo.
-
Todos los hijos de un hombre afectado serán afectados.
-
Cuando cruzaba plantas femeninas blancas con plantas verdes masculinas el resultado era blanco.
-
Cuando cruzaba plantas femeninas verdes con plantas masculinas blancas el resultado era verde.
-
Cuando cruzaba plantas femeninas verdes con plantas masculinas con fenotipo variegado el resultado era verde.
-
Cuando cruzaba plantas femeninas con fenotipo variegado con plantas masculinas verdes el resultado era variegado.
-
La mas común era un reparto equitativo de los cloroplastos con pigmento y sin pigmento.
-
La otra era un reparto no equitativo, es decir una se lleva los cloroplastos pigmentados y la otra los cloroplastos sin pigmentación.
-
Siempre se transmite a través de los gametos femeninos.
-
La heredan ambos sexos.
-
No hay salto de generaciones.
-
Edad: la calvicie, la enfermedad de huntington.
-
Influencia del sexo: puede ser de dos tipos.
-
Afecta de acuerdo con cual sea el sexo del cromosoma donador de un gen concreto.
-
Si un gen no se expresa porque lo ha recibido en el gamento de su madre. Es que ese gen está sometido a impronta materna.
-
Si un gen no se exprese porque lo ha recibido den el gameto de su padre. Es que ese gen está sometido a impronta paterna.
-
Dependiendo de cual sea el sexo de un organismo portador de un gen o genotipo concreto. Puede ser no al azar y puede ser al azar.
-
No al azar: el gen es responsable de la incontinencia pigmentaria. Esta ligado al X. Es letal y la letalidad solo se expresa en el sexo masculino.
-
Al azar: Mary Lyón en los sesenta lanzo la hipótesis de Lyón o lyonización “en todas las hembras de mamífero y como un mecanismo de compensación de la dosis genética se produce lo siguiente:
-
Uno de los cromosomas X se inactiva.
-
La inactivación se produce en las primeras etapas del desarrollo embrionario.
-
Una vez producida la inactivación del cromosoma X todas las células derivadas por mitosis de aquella que se inactivado un cromosoma X llevaran ese mismo cromosoma inactivo. (la inactivación se mantiene).”
-
No todos los genes del cromosoma X inactivo se inactivan porque hay genes que escapan a la inactivación.
-
Se sabe que en el brazo largo del cromosoma X concretamente en el XQ13 hay una región o centro llamado Xic que significa “centro de inactividad cromosómica”. En este locus cromosómico está situado un gen llamado XIST que solamente es activo en el cromosoma X inactivo y parece ser el responsable de la metilación y desmetilación reversible de los genes situados en el cromosoma X que se inactiva.
-
A pesar de que la hipótesis de Lyón se propuso como un mecanismo de compensación de la dosis génica y por tanto solo se aplico a las hembras que tienen dos cromosomas X ( por tanto doble dosis génica) y no se aplico a los hombres, se sabe que hay genes en el cromosoma Y que también se inactivan.
-
Si un homocigoto ChCh o un heterocigoto Chc se le cría a 30º todo su pelaje es albino.
-
Si se le cría a 25º aparece el fenotipo himalaya porque las zonas distales están a menor temperatura.
-
Si se le cría a 25º y se le pone un foco a una temperatura meno de 25º aparece pigmento.
-
norma de reacción: conjunto de fenotipos que es capaz de producir un genotipo concreto como consecuencia de la influencia de el fondo genético, factores ambientales.
-
Ruido de desarrollo: son los factores al azar que pueden influir sobre un genotipo y además son compatibles.
-
Epistasia simple y recesiva: en el color del pelo de las ratas. Dos fenotipos, se denomina simple porque solo ejerce epistasia 1 gen, y recesiva porque ese gene ejerce la epistasia en Homocigosis recesiva. caca impide la expresión del otro. Es decir no se da la pigmentación. Las relaciones mendelianas quedan 9:3:4
-
Epistasia simple y dominante: en el color del fruto de la calabaza. Dos fenotipos simple porque solo ejerce epistasia un gen y dominante porque ese gen la ejerce tanto en Homocigosis como en Heterocigosis. W en Heterocigosis impide la expresión de los otros genes. 12:3:1
-
Epistasia doble o complementaria: las dos formas de dominancia se complementan para producir el color son los genes no epistásicos. En el color de la flor del guisante hay dos fenotipos doble es porque la epistasia la ejercen dos genes y recesivas porque ambas genes la ejercen en Homocigosis recesiva. 9:7
-
Epistasia doble y dominante y recesiva o supresión: uno de los genes suprime el efecto del otro: en el genero primula la producción de malvidina. Dos fenotipos; doble porque son dos genes los que actúan y dominancia y recesiva porque uno de ellos es en Homocigosis y Heterocigosis y el otro solo en Homocigosis recesiva.13:3
-
Epistasia doble y dominante o duplicación se da cuando un gen en lugar solo en un locus esta en dos: en la bolsa de pastor el color del fruto. Dos fenotipos dos genes ejercen la epistasia y ambos en Heterocigosis y Homocigosis. Se le llama A1 y A2 porque es el mismo gen, pero repetido. 15:1
-
la mediada de F1 es aproximadamente intermedia entre las medias de las dos líneas puras parentales.
-
La medida de F2 es aproximadamente igual a la mediada de F1.
-
La F2 muestra mayor variabilidad alrededor de la media.
-
Los valores extremos de F2 se extienden mas que los valores extremos de F1.
-
Los caracteres cuantitativos están determinado por múltiples genes, se dice que están bajo control poligenico, a cada uno de los genes se le llama poligen o gen menor.
-
En general los caracteres bajo control poligenico se pueden cuantificar midiendo, pesando, contando etc.
-
Entre los alelos de un gen no hay dominancias, en lugar de hay alelos que contribuyen y hay alelos que no contribuyen a la expresión de un carácter.
-
Cada alelo que contribuye tiene igual efecto.
-
Los efectos de cada alelo que contribuye son aditivos.
-
No hay epistasias entre los genes no alelicos, es decir, entre genes diferentes.
-
No hay ligamiento, por tanto hay segregación y combinaciones independientes.
-
No hay efectos ambientales.
Prueba de alelismo ! análisis de complementación.
Cuando en una clonación aparecen fenotipos mutantes “variantes fenotípicos no usuales en la población” el investigador cuando va a iniciar el origen de los mismo, primero se plantea si surgen como consecuencia de que hay varios alelos de un gen o surgen como consecuencia de mutaciones en distintos genes.
Una manera de analizarlo es mediante una prueba de alelismo denominada análisis de complementación; no es la única, es una prueba para analizar lo que en esta parte de la genética (observación) esta es observado.
Definición “complementación”: se llamo complementación al intercambio de sustancias génicas (intercambio de la sustancia expresada por un gen).
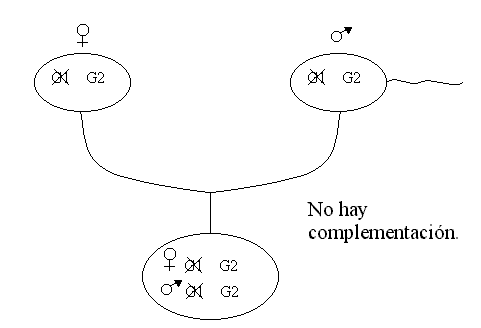
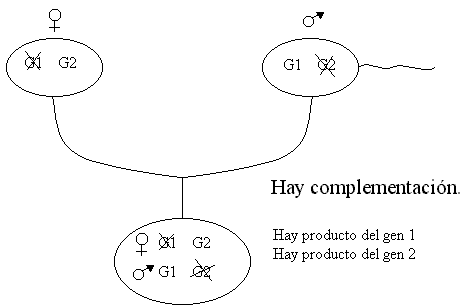
Si hay complementación deduzco que las mutaciones afectan a genes diferentes.
Si no hay complementación deduzco que las mutaciones afectan al mismo gen y por tanto generan alelos distintos del mismo gen.
Experimentos.
Conclusión final: algunas de las mutaciones que afectan al tamaño de las alas esta producidas por alelos de un mimo gen y sin embargo otras mutaciones que afectan al tamaño de las alas están producidas por mutaciones en genes diferentas.
A partir de esto se puede dibujar un esquema.
Un gen ! un carácter.
Un gen puede dar a mas de un carácter.
Cuenot se dio cuenta de esto en 1904. lo hizo con ratones mus musculus. Analizando los distintos fenotipos para ver si esto era concordante con lo dicho por Mendel.
Se encontró con que la mayoría tiene un color llamado agutí (negro jaspeado). Aquel gen lo denomino A luego vio que apareció una variante que era negro (todo el pelo pigmentado). Vio que el Agutí era dominante total sobre el negro. Al seguir cruzando ratones encontró un ratón amarillo.
Obtuvo líneas puras de Agutí, luego Amarillo con amarillo y obtuvo ratones amarillos y agutí en una relación de 2/3 : 1/3 Amarillo : agutí.
Conclusiones:
Para comprobar las conclusiones realiza el cruce prueba amarillo x agutí y le sale una relación de uno a uno de agutí con respecto al amarillo.
Cuando mira el útero de las ratoncitas amarillas descubre algunos ratoncillos amarillos muertos. Ese es el ¼ de homocigotos dominantes que faltaban. De ahí dedujo que: hay un alelo que llamó AY. Este alelo es responsable de dos caracteres “color amarillo” y “letalidad”.
Para el carácter amarillo basta con AY __ pero para el letalidad tiene que ser AY AY. Esto hace que los conceptos de dominancia y recesividad son atribuibles a los caracteres y no atribuibles a los genes. El color amarillo es dominante sobre el agutí y esta determinado por el gen AY, y el carácter letalidad es recesivo con respecto al carácter supervivencia y también esta determinado por el gen AY.
Falta un cacho del 16-10-2002
Fundamentalmente los genes que porta el cromosoma Y están relacionados con el sexo.
El SRY que es el que determina el sexo masculino, es esta zona mas importante de la determinación del sexo masculino en el hombre. Tiene un factor que es el TDF (factor que determina los testículos) pero en el hombre se le denomina SRY.
En Drosophila la presencia de dos cromosomas X determinan el sexo femenino, porque es un balance entre el numero de cromosomas autosómicos y el numero de cromosomas X. Si esta en pares la relación entre X y autosómicos es " 1 el sexo es hembra. Sin embargo cuando esa relacione es " 0.5 el sexo es masculino.
Tipos de ligamiento a sexo.
Dos tipos: total o parcial.
Los genes situados en los cromosomas sexuales y que no tienen homología entre ellos, muestran un tipo de herencia con ligamiento total al sexo. Coso del gen W responsable del color de los ojos en Drosophila.
Los genes situados en los cromosomas sexuales y que tienen homología entre ellos, se dice que tienen un tipo de herencia parcialmente ligados al sexo.
Herencia influida y herencia limitada por el sexo.
Algunos caracteres como por ejemplo los cuernos en vacuno o la calvicie en humanos esta influenciados por el sexo y esta influencia se lleva a cabo a través de hormonas sexuales.
| Genotipo | Fenotipo. | |
| Femenino | Masculino | |
| BB | Calvo | Calvo |
| Bb | No calvo | Clavo |
| bb | No calvo | No calvo |
La aparición de este fenómeno esta determinado por un gen en dos formas alelicas H, h. La forma H inhibe la formación del plumaje masculino y además en presencia de cualquier tipo de hormona. Pero cuando esta presente la forma h no se inhibe la formación del plumaje masculino pero este no puede aparecer en presencia de hormonas femeninas.
| Genotipo | Fenotipo | |
| Femenino | Masculino | |
| HH | Plumaje gallina | Plumaje gallina |
| Hh | Plumaje gallina | Plumaje gallina |
| hh | Plumaje gallina | Plumaje gallo |
Análisis de árboles genealógicos de cromosomas sexuales.
X dominante.
X recesivo.
Ligado al Y.
Herencia extranuclear.
Los genes que Mendel analizo estaban en el núcleo de la célula pero existen también genes situados en orgánulos citoplasmáticos. Estos son los responsables de la herencia extranuclear y no sigue el comportamiento de Mendel.
El investigador que los descubrió fue Correns en 1909. el organismo a estudio fue Mirabilis Jalapa. Lo que Correns estudió es la herencia del color de las hojas. En términos generales el color de las hojas es verde, pero con los entrecruzamientos Correns se encontró con plantas que con respecto a este fenotipo presentaban variantes a lo que es usual. Encontró que aparecían plantas con algunas hojas de color verde, otras hojas de color blanco y otras hojas de color verde y blanco, es decir había un fenotipo variegado (sin distribución uniforme). Se pregunto el origen, y comenzó a hacer cruzamientos, así encontró lo siguiente.
Conclusión: cuando cruzaba plantas los descendientes siempre mostraban el fenotipo de la parte femenina de la planta (planta madre).
Debido a esto Corres pensó que si el gamento femenino tiene núcleo y citoplasma y el masculino solo núcleo estos genes estaban en el citoplasma.
Correns propuso que esto está en genes situados en los cloroplastos.
Para explicar porque aparecían hojas blancas, verdes y con manchas, él propuso que las células que van a dar ese fenotipo variegado tiene que tener dos tipos de cloroplastos, tiene que tener cloroplastos con pigmento verde y cloroplastos no pigmentados.
Cuando se produce la citocinesis se produce también la segregación citoplasmática. Es decir, la separación de todas las moléculas, orgánulos, etc que hay en el citoplasma, y esta segregación se podía producir de dos maneras.
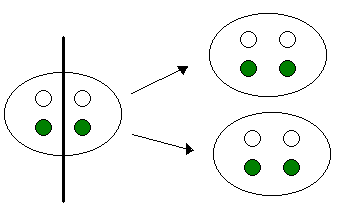
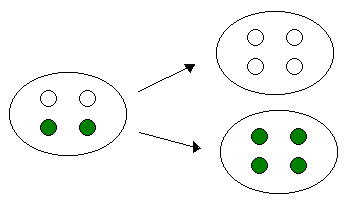
CARACTERÍSTICAS DE LA HERENCIA EXTRANUCLEAR.
Genes ! fenotipo (no univoca).
Relación univoca: Siempre que hay un genotipo se le asocia un fenotipo concreto
ii! 0 (invariablemente)
IAIB ! AB (invariablemente)
Factores que influyen en los genes.
Es importante decir que los factores no los mutan ni los modifican pero intervienen en la expresión del gen.
Entre los factores hay de dos tipos:
1.- Fondo genético.
Un gen concreto ocupa habitualmente un locus cromosómico, el entorno genético que lo rodea es el fondo genético y eso tiene influencia en la expresión del gen.
Si el gen W+ que ocupa un locus especifico en el cromosoma salta y pasa a una zona heterocromática se inactiva. Esto es el efecto posicional; la expresión de un gen puede verse afectada por la posición en el genoma. No ha mutado, solo ha cambiado el locus génico.
A todos los mecanismos que influyen en las expresión de los genes sin modificar su secuencia y además son transmisibles son conocidos como mecanismos epigenéticos y la herencia es: herencia epigenética. De los mecanismos epigenéticos que dan lugar a la herencia epigenética el mas conocido es la metilación.
Metilación: adición de un grupo metilo a las bases nitrogenadas del DNA, y la base nitrogenada que preferentemente puede metilarse es la citosina, y la posición en la cual se metila es la posición 5'. Preferentemente en las islas CPS. La metilación genera inactividad génica porque contribuye al empaquetamiento de la cromatina y sin embargo la desmetilación aumenta o contribuye al aumento de la actividad génica porque potencia el desempaquetamiento del DNA (cromatina).
2.- Factores ambientales:
A) AMBIENTE INTERNO:
Hay dos: la influencia de la edad y la influencia del sexo.
Ejemplo: existe un fenomenito denominado Impronta geonómico o impronta parental (del ingles imprinting) descubierto hace muy poco:
Hay genes que se expresan o no dependiendo de que sea el padre o la madre el que a transmitido ese gen al hijo.
Ejemplo de impronta materna: ratones enanos. El gen esta sometido a impronta materna y cuando el ratoncillo lo recibe no se expresa y si del padre ha recibido el gen mutado el ratón sale enano.
Este fenómeno de impronta esta producido por un mecanismo epigenético, es un mecanismo que es hereditario sin que el mecanismo afecte a la secuencia del gen, es decir solo afecta a su expresión.
Muchos de los procesos producidos son de metilación, y son reversibles.
El numero de cromosomas X inactivados es igual a el numero de cromosomas X que se posean menos uno.
A nivel citogenético en la parte interna y pegado a la parte interna de la membrana nuclear. Cuando se tiñen los núcleos de las hembras normalmente de mamíferos aparece una manchita intensamente teñida y se denomina corpúsculo de Barr (ya que fue este investigador quien lo descubrió); es un cromosoma X inactivo.
El corpúsculo de Barr es heterocromatina pero no siempre es fija en unas células esta en un sitio y en otras en otro sitio distinto por ello se le denomina heterocromatina facultativa dado que cambia de lugar según la célula observada.
La lyonizacion es un mecanismo epigenético, en este caso también parece estar implicada la metilación.
A la hipótesis de Lyón podemos añadirle:
B) FACTORES AMBIENTALES EXTERNOS.
1) De naturaleza física (temperatura).
En ciertos mamíferos (gatos, ratones y conejos) respecto al fenotipo “color del pelaje” hay uno denominado Himalaya que es que todas las partes son albinas a excepción de ciertas partes que tienen temperatura mas baja como son las patas, la cola, las orejas y el hocico.
Tabla 4,2.
Hay cuatro formas alelicas C> Cch >Ch > c. Entre ellos hay uno Ch, es el gen cuya expresión se ve influido por la temperatura.
2) De naturaleza Química (medicamentos).
En los años 50-60, a las embarazadas se les daba una medicina llamada talidomida para paliar los síntomas como nauseas, vómitos, etc. Se administraba en los primeros tres meses de embarazo cuando el bebe estaba aun en formación, estos niños nacían con grandísimos defectos en el desarrollo de las extremidades porque influía en la expresión de un gen.
Consecuencias.
Variable expresividad.
Como consecuencia del efecto que los diferentes factores tienen en la expresión génica. Tiene una variable expresividad, entendiendo por esto “al grado de intensidad en que un genotipo se expresa en un fenotipo”.
Ejemplo: osteogenesis imperfecta.
Aquí vemos como no afecta de igual grado a todos los miembros de la familia afectado, dado que los síntomas de esta enfermedad es huesos de cristal, sordera y problemas de visión. Pueden darse individuos que solo tengan uno, dos o todos los síntomas.
Variable penetrancia.
Es el “numero de individuos que expresan el fenotipo asociado a un genotipo” puede suceder que alguien teniendo un genotipo concreto no exprese el fenotipo.
Mas fenotipos.
Un genotipo concreto da lugar a múltiples fenotipos como consecuencia de la influencia ambiental.
Fig. 1.2
Fenocopias.
Cuando un efecto ambienta produce un fenotipo que es una copia o una imitación del fenotipo producido por un genotipo concreto al primero de le denomina fenocopia.
Ejemplo: Talidomida.
Es un efecto ambiental que produce un fenotipo “alteraciones del desarrollo de las extremidades” pero este fenotipo es una copia de fenotipo producido por un gen que cuando muta produce el síndrome de Oram-Holt.
Varios genes ! un carácter.
A diferencia de Mendel hoy se sabe que varios genes pueden intervenir en la manifestación de un carácter este hecho determina que ese carácter aparezca en muchas formas fenotípicas diferentes. Estas formas fenotípicas alternativas pueden mostrar un variación discontinua y además pueden mostrar una variación continua.
Variación discontinua.
Los diferentes genes interactúan entre ellos en la expresión de un fenotipo pero sin epistasias, o pueden generar epistasias, lo cual implica que alguno de los genes puede inhibir la expresión de otro u otros genes; estos son los genes epistáticos. Aquellos genes a los que modifican o inhiben son los genes hipostáticos.
! Sin epistasias: cuando dos o mas genes se combinan la consecuencia es la aparición de mas fenotipos. Pueden darse dos alternativas, que no se modifiquen las relaciones mendelianas digenicas. O que si se modifiquen las relaciones mendelianas digenicas.
Ejemplo: la forma de la cresta de las gallinas. Se dan 4 alternativas producidas por dos genes P y R cuando los dos genes se juntan desde el punto de vista genotípico hay 4 combinaciones. Cada una de esta produce un fenotipo diferente. Y no hay modificación de las relaciones mendelianas digenicas.
! Con epistasias. Cuando uno o alguno de los genes que intervienen en la manifestación de un carácter dificulta o impide la expresión de otro y de otros genes que intervienen en la manifestación de ese carácter.
Ejemplo: la forma de la flor de la calabaza. Se produce cuando los dos heterocigotos producen un mismo fenotipo. Hay modificación de las relaciones mendelianas digenicas.
Siempre se produce una alteración de las proporciones mendelianas vigentes. Hay varios tipos y cada uno de ellos se reconoce por las diferentes alteraciones:
Variación continua.
Varios genes en manifestación de un carácter en un numero de individuos grande. Deben describirse en términos cuantitativos, a su estudio se le llama estudio de la herencia cuantitativa.
Los investigadores fueron Emerson y East en 1913. Utilizaron el maíz (Zea mays) y estudiaron la longitud de la mazorca. Siguiendo la metodología de Mendel cruzaron líneas puras en relación a las longitudes de l mazorca de maíz. Unas líneas eran cortas y otras largas.
Los descendientes tuvieron una longitud media de líneas puras y cortas; la longitud media de corto era " 6.66cm de longitud, la longitud media de largo era " 16.8 cm, la longitud media de las mazorcas en la F1 era " 12. 12 cm. Además al cruzar las plantas de F1 uno obtenían un F2 con una longitud media de 12.85 cm, había mas variación de longitud en relación a la F1.
Esto revelaron 4 datos, aplicarse a los siguientes rasgos que muestran una herencia cuantitativa:
Galton y Pearson (finales del 19) peso, color... demuestran cierta coincidencia entre padre e hijo.
Johannsen (1903) herencia del peso de las judías: variación del peso con base genética.
Wilson-Ehle (1909) la variación continua estaba determinada por múltiples genes, y que cada uno de estos segrega de acuerdo con los principios mendelianos.
Hipótesis poligenica o herencia poligenica.
Fue propuesta por Nilson L., hipótesis poligenica o de caracteres múltiples.
22
La relacione entre IAi y IBi es una relación de dominancia total, en ambos casos el heterocigoto manifiesta un solo fenotipo el del alelo dominante.
La relación entre los dos alelos es de codominancia ambos se manifiestan
Descargar
| Enviado por: | Amae |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.