Ingeniero Químico
Transporte de iones a través de canales en membrana celular
TRANSPORTE DE IONES A TRAVÉS DE CANALES EN LA MEMBRANA CELULAR
INTRODUCCIÓN
Los cien mil millones de células del cuerpo humano no podrían existir si cada una de ellas no estuviera rodeada de una membrana que la separa de sus vecinas y del líquido exterior. Tanto dentro como fuera de la célula el agua constituye el disolvente y el vehículo por el que moléculas como aminoácidos, proteínas, o el mismo ADN permanecen disueltos en ella y realizan sus funciones, actuando las membranas a modo de barreras de tipo graso que impiden que las moléculas circulen anárquicamente por todo el organismo. Sin embargo, las células necesitan comunicarse constantemente entre sí, a través de señales que atraviesan sus membranas, y que no son más que pequeñas moléculas o iones que mediante una cascada de reacciones químicas permiten que las funciones biológicas tengan lugar.
Roderick MacKinnon, obtuvo importantes hallazgos sobre el canal de potasio y de otros iones, como los cloruros. Los canales iónicos regulan, entre otras, las funciones del sistema nervioso y de los músculos. En el impulso nervioso, un canal iónico se abre en la superficie de una neurona como respuesta a una señal química emitida por otra neurona vecina, y el pulso eléctrico así generado se propaga mediante la apertura y cierre de otros canales, hasta la neurona siguiente, y todo ello en unos pocos milisegundos.
Siempre ha resultado intrigante la selectividad de los canales de iones. Tanto el sodio como el potasio son cationes esféricos con carga +1 y diámetros de 1,90 y 2,60 ángstrom, respectivamente. El sodio es, por tanto, algo menor. ¿Cómo explicar la selectividad del canal de potasio, que no deja pasar al sodio, un ión más pequeño?
DESARROLLO
El canal de potasio.
Estos canales son proteínas que se encuentran en todas las células que posean membrana. No hay forma en que los iones atraviesen la membrana si no es a través de canales.
La membrana celular está formada por lípidos (grasas), y las sales, que no se mezclan con las grasas, no pueden atravesar esa pared que protege a la célula. Por consiguiente, la entrada sólo puede realizarse a través de canales.
Los canales iónicos no son estructuras que están abiertas todo el tiempo, sino que se abren y se cierran de acuerdo con las órdenes que reciban, y esa apertura y cierre puede modularse. De hecho, tienen la ventaja de responder a estímulos de manera muy rápida, en menos de milisegundos.
MacKinnon estudió, en particular, el canal de potasio, que se puede modular mediante cambios de voltaje. Estos canales de potasio ayudan a propagar los impulsos eléctricos en el cerebro y el corazón.
El movimiento de los iones se debe fundamentalmente a dos efectos:
-
Difusión: en presencia de un gradiente de concentración (provoca la entrada de iones de potasio al interior de la célula)
-
Atracción eléctrica: en presencia de un campo eléctrico (provoca la salida de iones de potasio del interior de la célula al exterior)
Cuando un impulso eléctrico viaja a lo largo de un nervio, la carga a lo largo de la membrana celular se modifica y el exterior de la membrana se carga negativamente con respecto al interior. Este cambio en la carga hace que los canales de potasio se habrán y permitan que los iones de potasio salgan de la célula.
La salida de potasio hace que la membrana se encuentre cargada negativamente, contrarrestándolo por difusión y volviendo a su estado de reposo (en condiciones normales, en el interior de la célula hay una concentración 20 veces superior que en el exterior de iones de potasio) preparándose así para el siguiente impulso nervioso.
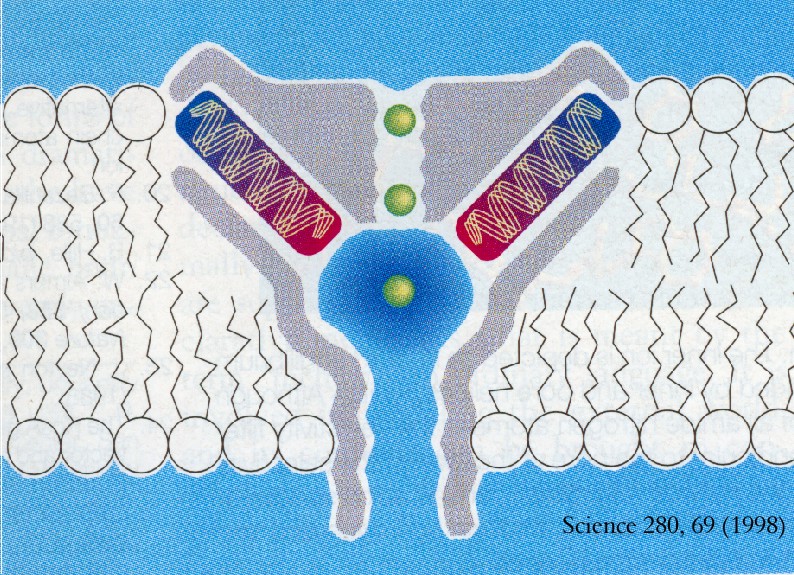
Estructura del poro, y función de las partes que lo componen.
El canal de potasio es un tetrámero, formado por cuatro subunidades idénticas. Las cuatro subunidades se insertan en la membrana dejando un poro en el interior.
El poro tiene una longitud de 45 Å y su diámetro varía a lo largo de éste. Podemos dividir al poro en tres regiones, desde dentro: un túnel de 18 Å cuyas paredes lo constituyen las hélices internas, una cavidad acuosa a mitad de la membrana y una región más estrecha flanqueada por los filtros de selectividad que separan al poro de la solución extracelular.
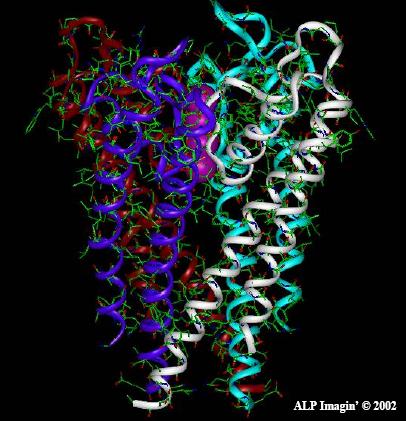
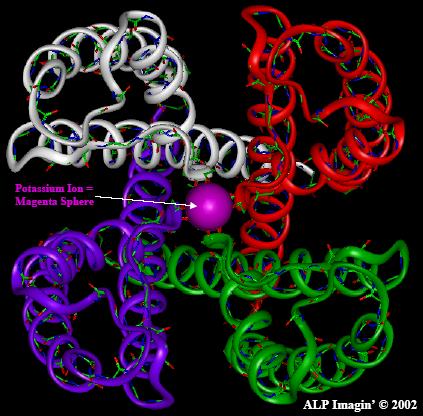
Las figuras superiores muestran las vistas en planta y alzado respectivamente de las hélices que componen el poro.
Para el paso selectivo de iones este canal dispone de dos mecanismos:
El de la compuerta (que se abre o cierra por estímulos del entorno, ya que una de las “colas” del tetrámero se desliza sobre el poro y lo cierra).
El del filtro, que solo permite el paso de una molécula de sodio por cada diez mil de potasio, lo que lo hace tremendamente selectivo.
Mecanismo de filtro
La cavidad acuosa se forma cuando un catión crea un campo electrostático que polariza su entorno, atrayendo los polos negativos de los dipolos hacia él. En una bicapa lipídica la polaridad es nula y por esto la energía del catión es muy alta. Para contrarrestar esta energía existe la cavidad acuosa.
En solución, los iones están estabilizados por moléculas de agua que orientan sus átomos de oxígeno alrededor de la esfera cargada del catión. Las hélices se orientan de manera que el extremo amino (+) queda alejado de la cavidad y su extremo carboxilo (-) cercano a ésta donde está alojado el catión. A esto se le conoce como efecto hélice dipolo, es otra estrategia para compensar la energía del catión en el interior de la membrana.
En el interior del canal, una serie de átomos de oxígeno de los grupos carboxilos se encuentran colocados exactamente igual que los del agua alrededor del potasio antes de entrar en el tubo, por lo que despojarse de su capa de hidratación no le supone al potasio gasto alguno en energía.
El radio atómico del potasio es 1.33 Å y el del sodio es 0.95 Å, con solamente esta diferencia: los canales de potasio manejan la selección para el ión potasio sobre el ión de sodio por un factor de más de mil. El sodio, al perder las moléculas de agua que lo estabilizan, quedaría dentro del canal en una situación desfavorable, con los oxígenos demasiado alejados. Podríamos decir que bailaría dentro del tubo sin ajustarse perfectamente al mismo, por lo que prefiere quedarse fuera.
Por su alta selectividad, se pensaría que el potasio quedaría firmemente unido dentro del canal y no difundiera rápidamente por el poro, pero esto no suele ocurrir por dos razones:
a.- Primero el filtro de selectividad contiene más de un ión de modo que la repulsión entre los iones que están cerca ayudan a superar la afinidad intrínseca que cada ión tiene para su sitio obligatorio.
b.- La estructura del filtro de selectividad depende de la presencia de los iones de potasio. La entrada del segundo ión hace una cambio conformacional del filtro, lo cual hace que los demás iones se aten menos firme y difundan mas fácilmente. La conformación conductora del filtro requiere la presencia de dos iones de potasio y es el segundo el que induce el cambio conformacional del canal.
En conclusión, el canal resulta estar tan bien adaptado a los iones de potasio que casi no necesitan energía para atravesarlo, excluyendo de esa forma otros iones, que no tienen suficiente energía para atravesar el canal. Esto explica cómo pueden ser tan rápidos y al mismo tiempo tan selectivos.
El posicionamiento de las hélices de la proteína, explica que los iones atraviesen la membrana en una dirección pero no en la contraria, y la presencia de una bolsa o piscina de agua en medio de la membrana, ayuda a que el viaje del ión sea más corto.
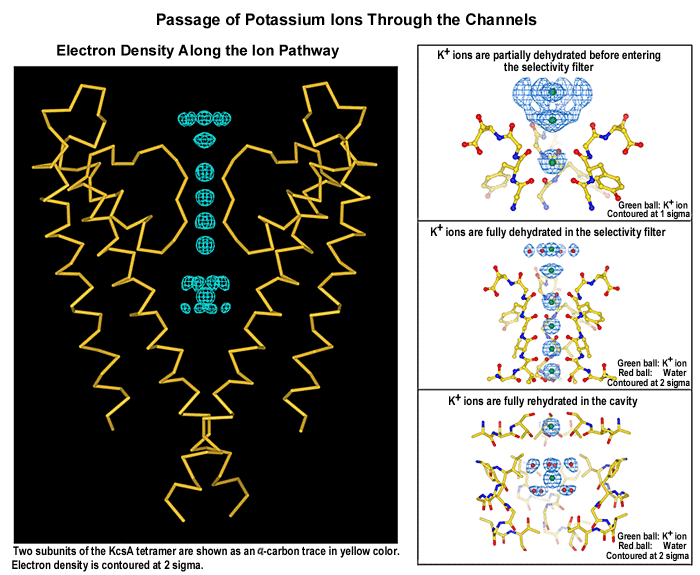
Mecanismo de compuerta
Una vez que se abre el canal, debe existir un mecanismo para cerrarlo. El mecanismo mas aceptado es el de la cadena y la bola. Según este modelo una compuerta del lado extracitoplasmático se cierra en ausencia de iones de potasio provenientes del exterior, Esta compuerta suelen ser los primeros residuos de aminoácidos de una subunidad alfa o beta (o ambas). Un canal iónico se puede volver a abrir, por lo que el canal, al cerrarse, no forma ningún enlace covalente nuevo. El sistema de cerrar las compuertas está relacionado con el desfavorecer el flujo del ión en cierta dirección. Cuando la concentración del ión cambia, también ocurre un cambio en el potencial de Nernst para este ión. Esto causa el cambio conformacional. Sin embargo, al menos en los canales de potasio operados por voltaje, este no es el único mecanismo para cerrar el canal.
El otro mecanismo parece no ser tan usado por la célula. Este se basa en un ión de magnesio que tapa el canal. El magnesio se encuentra en el citoplasma, y en ciertas condiciones se ve atraído al canal. Sin embargo, si esto pasa se tiene resultado, cerrar la compuerta.
CONCLUSIÓN
Estudios realizados por investigadores del Instituto Médico Howard Hughes han identificado la forma en la que los canales de potasio se abren para permitir que los iones potasio atraviesen la membrana celular —proceso que es crucial en muchos procesos biológicos, incluyendo el latido rítmico del corazón y la generación de los impulsos nerviosos—.
Las alteraciones en la función del canal de potasio también se han relacionado con la diabetes, enfermedades cardíacas, asma y otras enfermedades. La comprensión de los principales detalles de la forma en la que funcionan estos canales eventualmente podría ayudar en el diseño de nuevas drogas para tratar distintos trastornos.
BIBLIOGRAFÍA:
http://www.elpais.es/articulo/elpfutpor
http://www.hhmi.org/news/mackinnon9-esp.html
http://personales.com/chile/valdivia/canalesdepotasio/
http://www.hhmi.org/news/mackinnon6-esp.html
6
Descargar
| Enviado por: | SiLvA |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.