Psicología
Psicología fisiológica
TEMA 2: TÉCNICAS EN PSICOLOGÍA FISIOLÓGICA
Introducción al aparato estereotáxico
-
Método de las operaciones convergentes: llegar a los mismos resultados a través de diferentes técnicas de experimentación.
-
Aparato estereotáxico: dispositivo mecánico capaz de inmovilizar la cabeza del sujeto experimental anestesiado y localizar cualquier estructura cerebral, con precisión milimétrica.
-
Partes: barras auditivas, pinza, portaelectrodos.
-
Escalas en las que se mueve el portaelectrodos en 3D: antero-posterior/lateral-medial/dorsal-ventral
Atlas anatómico
Consiste en una serie de dibujos o fotos que corresponden a las diferentes secciones cerebrales, parte de la premisa de que como no hay dos cerebros iguales podemos localizar cualquier estructura partiendo de características externas. Tomando como referencia tres puntos: alfa, beta e interaural.
Técnicas de supresión de actividad.
ELÉCTRICAS
Técnicas de lesión electrolítica: es una de las más usadas, se coloca por medio del aparato estereotáxico un electrodo de acero y oro en el interior del cerebro, con un generador se provoca una descarga de corriente eléctrica continua.
Vse puede controlar el tamaño de la lesión, no se dañan las zonas adyacentes.
Ila corriente destruye por igual cuerpos celulares y fibras, si no controlamos los parámetros puede afectar a zonas adyacentes.
Técnicas de radiofrecuencia: el daño se produce por el calor producido por la resistencia que impone al tejido cerebral al paso de corriente eléctrica alterna que suministra un electrodo.
Ies menos específica que la electrolítica, no se controla la difusión del calor y se dañan zonas adyacentes, solo se puede usar superficialmente.
Técnicas de electrocauterizacion: Se calienta una micropipeta de vidrio con corriente eléctrica y se introduce en el cerebro, causando la lesión ( no es calor resultante)
Isolo se usa superficialmente
MECÁNICAS.
Técnicas de aspiración: con una pipeta conectada a un sistema de vacío se aspira el tejido que queremos lesionar.
Ies muy inespecífico y puede provocar infecciones graves.
Técnicas de ablación (lobotomía): mediante un microbisturí se extirpan las zonas que queremos lesionar.
Ies muy imprecisa.
Técnicas de sección: se coloca un microbisturía (knife-cut) en el aparato estereotáxico y se seccionan las fibras que queremos lesionar.
Ves mas específica que las anteriores.
QUÍMICAS E IRREVERSIBLES.
Neurotoxinas: mediante sustancias como el ácido iboténico y el ácido kaínico se produce la destrucción selectiva de cuerpos celulares dejando intactos los axones.
Ino es de las mas específicas.
Inactivadores de los neurotransmisores:
-
Inactivadores del sistema dopaminérgico: sustancias como 6-OH-DA destruyen el sistema catecolaminércigo (adrenalina, noradrenalina, dopamina)
-
Inactivadores del sistema serotoninérgico: sustancias como 5-7-DOH-5-HT inactivan las neuronas serotoninérgicas.
-
Inactivadores del sistema colinérgico: sustancias como AF-64 destruyen las neuronas colinérgicas.
Vson técnicas mas específicas que las anteriores.
Ise administran en suspensión, pueden afectar a zonas adyacentes, y todas son irreversibles.
QUÍMICAS Y REVERSIBLES.
Anestésicos locales: sustancias como la lidocaína, procaina y novocaína inactivan temporalmente la zona a manipular, bloqueando la transmisión.
Vsu efecto aparece muy rápido. 10 seg.
Isu efecto es poco duradero (dos minutos)
Cationes: sustancias como el magnesio, cobalto o manganeso inactivan los canales de calcio, impidiendo que se produza el potencial de acción
Isu efecto es poco duradero, y si se administra repetidamente se produce muerte celular.
Agonistas de GABA: sustancias como el muscitol, inhiben la acción del GABA.
Vse puede administrar de forma repetida, pues no es tóxico.
Iduracion limitada, 90-100 seg.
TTX (tetradotoxina): bloquea los canales de sodio, impidiendo la propagación del potencial de acción.
V su efecto es duradero (110 min), puedo estudiar la función de una zona cerebral determinada, los datos son mas fiables porque el propio sujeto es el control, reduce el número de sujetos necesarios, puedo estudiar la implicación de varias zonas con el mismo sujeto.
Itarda 10 minutos en aparecer su efecto.
GENÉTICAS.
Knock-out: es la mutación de un gen particular.
Ipueden aparecer respuestas compensariorias de otros genes, y las funciones del gen desaparecen de todo el organismo y no de una zona.
CRIOGÉNICAS.
Técnica de inactivación criogénica: se basa en enfriar una zona cereberl con un liquido refrigerante.
Ves reversible.
Técnicas de estimulación cerebral.
Técnicas de estimulación eléctrica: se coloca un electrodo en el cerebro, el poco + en el interior y el polo - adosado al cráneo y se estimula la región de forma normal (fisiológica). Esta estimulación provoca una despolarización de las nueronas.
Vse pueden controlar los parámetros de estimulación: frecuencia, intensidad y duración, la estimulación es específica de la zona.
Técnicas electroconvulsivas. TEC: se colocan dos electrodos en las sienes y se aplica una descarga fuerte de breves segundos.
Ves útil en pacientes con depresión
Ipoco ortodoxa y con efectos secundarios.
Ténicas de estimulación química: consisten en la administración de sustancias químicas en el cerebro que activan neurotransmisores específicos.
Vdiferencia entre tipos de neuronas.
Ihay que esperar que el fármaco haga efecto, la difusión puede afectar a zonas adyacentes.
Técnicas de registro de actividad cerebral.
Electroencefalograma. EEG. Consiste en colocar unos electrodos en el cuero cabelludo y registrar la actividad en una amplia zona.
Vresolución temporal perfecta.
Iescasa resolución espacial
-
Registro de potenciales evocados: consiste en usar el EEG para registrar la actividad cerebral resultante ante la presentación de un estímulo.
Ise necesita eliminar la actividad cerebral espontánea
-
Registros promediados evocados: resultados de un programa de ordenador que regrista los potenciales evocados quitando la actividad espontánea.
Registros multicelulares: consiste en electrodos que registran la actividad de las neuronas que cae bajo su radio de acción.
Vmejor resolución espacial que el EEG
Ino diferencia entre tipos de neuronas.
Registros unicelulares: consiste en implantar unos electrodos muy finos en el cerebro del animal que registra la actividad desde el espacio extracelular.
Técnicas de neuroimagen.
Técnicas de neuroimagen estáticas.
Angiografía cerebral: se administra un colorante radio-opaco vía intravenosa y luego mediante rayos X se puede ver el sistema circulatorio.
Vutil para tumores
Ino diferencia entre estructuras cerebrales.
Pneumoencefalografía: permite ver el sistema ventricular. Se extrae el LCR y se sustituye por aire.
Tomografía axial computarizada. TAC: se basa en la absorción de rayos X por los tejidos, permite diferenciar la sustancia blanca de la gris.
Vbuena resolución temporal.
Isolo ofrece fotos de cortes basales u horizontales. No se puede aplicar de forma repetida. No informa del funcionamiento.
Resonancia magnética nuclear. RMN: Se basa en la detección de protones y funcionan como campos magnéticos. Estos son captados por potentes imanes que mediante complejos programas de ordenador producen imágenes en 3D.
Técnicas de neuroimagen dinámicas.
Autoradiografía: se basa en el consumo de glucosa por el organismo, por tanto, sustituyendo la glucosa por un análogo que no puede ser degradado, este se queda en el lugar demandado. Si además marcamos radioactivamente la podremos detectar mejor.
Vaporta resolución espacial y temporal.
Tomografía computerizada por emisión de un simple fotón: se basa en la detección de cambios en el flujo sanguíneo de determinadas regiones, para ello se inyectan productos radioactivos, se colocan unas cámaras alrededor de la cabeza y proporcionan imágenes en 3D en el ordenador.
Tomografía por emisión de positrones. PET: Basada en los principios del flujo cerebral. En este caso se inyectan productos que usa el organismo (oxígeno, nitrógeno, carbono…) pero con un marcaje radioactivo. Estos productos producen positrones que se autoaniquilan y provocan dos rayos alfa en direcciones opuestas que son captados por ordenadores donde aparecen las zonas mas activas en un color más nítido.
Resonancia magnética funcional. RMF.: se basa en la detección de los niveles de oxígeno.
Localización histológica: para descubrir que la manipulación se hizo correctamente o para descubrir las comunicaciones nerviosas. Fases.
-
Fase de perfusión: se anestesia al animal, se accede al ventrículo derecho por la caja torácica para eliminar la sangre y posteriormente se inyecta en el ventrículo izquierdo suero fisiológico isotónico para limpiar el cerebro. Finalmente se le inyecta una solución de formol al 10% para endurecer los tejidos.
-
Fase de extracción: se decapita al animal y se eliminan los tejidos que puedan impedir la extracción del cerebro. Se extrae y se deposita en un frasco con formol durante 24 horas.
-
Fase de laminación: se usa un microtorno. Esta compuesto por un sistema de calibración, una plataforma de congelación y una cuchilla afilada, se lamina el cerebro y se colocan las láminas en un portaobjetos y se cubre con un cubreobjetos.
-
Fase de tinción:
-
Tinción de golgi: diferencia las partes de las neuronas
-
Violeta de cresillo: revela los cuerpos celulares
-
Azul de metileno: revela la sustancia de Nills
-
Tinción de Weig- Weigert: revela las vainas de mielina.
-
Histología fluorescente: diferencia entre los sistemas de transmisión(azul: catecolamina, verde: serotonina)
-
Trazado de conexiones eferentes (sustancias de trazado anterógrado) como la lecitina PHA-L.
-
Trazado de conexiones aferentes: oro fluorado y peróxido de rábano
TEMA 3: EL DOLOR COMO PROCESO SOMATOSENSORIAL
Introducción
Factores diferenciadores del dolor con respecto a otros sistemas sensoriales
-
Se encuentra influenciado por factores psicológicos, su progresión se incrementa cuando ha llegado al umbral de detección.
-
Existen dos tipos, uno fásico o delta y otro crónico o tipo c. la existencia de estos dos tipos de dolor se manifiesta en el síndrome del doble dolor, pues se comprueba que cuando se recibe daño, primero se siente un dolor fásico que más tarde pasa a ser un dolor más intenso. Esto se debe a que el sustrato biológico de ambos es diferente. En el caso del dolor fásico, lo transmiten fibras mielinizadas, en el crónico lo procesan fibras c no mielinizadas.
-
No es percibido de la misma forma en los distintos momentos del día.
-
Dificultad para medirlo
-
Hay casos que no son tratables y otros que no han percibido dolor
Por todo esto el dolor no puede ser considerado como un sistema sensorial puro.
Bases psicobiológicas del dolor.
Mecanismos periféricos de procesamiento
-
El dolor se lleva a cabo por los mismos sistemas que procesan otras sensaciones, con el tiempo se demostró que no es cierto
-
Todos los sistemas sensoriales comparten el mismo sistema anatómico, tampoco resultó cierta
-
Una tercera propuesta ha confirmado que para el dolor existe un sustrato anatómico especializado, constituido por células pertenecientes a la categoría de terminaciones nerviosas libres.
Sistema nervioso central: vías y centros anatómicos del dolor.
Los cambios en el tejido producen la liberación de unas sustancias químicas (algógenas: producen dolor o prostangladinas: bajan el umbral del dolor).
Cuando todas estas sustancias actúan sobre la membrana celular (neurona de primer orden periférica) producen la apertura de canales iónicos, dando lugar a la entrada de iones + dando lugar a la despolarización del receptor y generando un potencial de membrana, que es graduado y se transfiere al soma de las neuronas que están en contacto con la piel (neuronas de primer orden) y que en encuentran en los ganglios dorsales de la médula espinal. El axón entra en la médula a través de la raíz dorsal, para hacer sinápsis con la neurona de segundo orden, que tiene su cuerpo celular en el asta dorsal de la médula, en la sustancia gelatinosa.
Estas neuronas cruzan al lado contrario, ocupando una posición ventrolateral, a partir de aquí se forma la vía principal del dolor, el tracto espinotalámico, que a su vez puede ser subidividido en dos partes, el compuesto por fibras delta (neoespinotalámico) y el compuesto por fibras neurales “c” (paleoestinotalámico)
-
A nivel del puente el tracto neoespinotalámico ocupa una posición lateral, a nivel del mesencéfalo en la zona ventrobasal del tálamo y en la corteza proyecta al cortex somatosensorial primario que a su vez envía información al córtex somatosensorial secundario.
-
La vía paleoespinotalámica, a nivel del puente ocupa una posición medial, que lleva a cabo varios relevos a nivel de la formación reticular, el núcleo parabraquial y el locus cerúleos (todas ellas en el tronco), posteriormente tiene su relevo en la sustancia gris central del mesencéfalo(PGA) el dolor se sitúa en los núcleos talámicos ventromedial, parafascicular e intralaminar. Cuando asciende a la corteza, se activan zonas de la corteza cinculada anterior (A24) (recibe información del tálamo medial y parece importante en los componentes afectivos)
Diferentes modalidades del control del dolor.
Teoría de la puerta de control.
El control del dolor se produce en la médula, concretamente en la sustancia gelatinosa, el proceso es el que sigue:
Cuando una persona siente dolor, este es captado por las fibras pequeñas (c o delta) que lo transfieren al cerebro, al mismo tiempo, estas neuronas envían una colateral que inhibe a la interneurona de la sustancia gelatinosa, al estar inhibida no puede ejercer su función y el dolor no puede ser bloqueado. Pero si al mismo tiempo que el sujeto siente dolor que recibe una sensación de tacto o presión, esta sensación es captada por las fibras G o gruesas, que transmiten por su vía principal la sensación de tacto o presión, al mismo tiempo que envían una colateral a la sustancia gelatinosa, activando a la neurona inhibitoria que interrumpe la sensación de dolor, ya que no es capaz de pasar a la célula de proyección.
Analgesia por estimulación cerebral
Reynolds fue capaz de intervenir sin anestesia, lo consiguió estimulando la sustancia gris central del mesencéfalo (PGA)
Su analgesia parece deberse a las proyecciones descendentes que se inician en este núcleo y que terminan en la médula espinal, que aunque hay conexiones directas con la médula, la mayoría pasan por otros núcleos, por ejemplo:
Una de las más importantes es la que se establece con el núcleo del Rafe Magnus, sus fibras establecen contactos con el asta dorsal de la médula espinal, inhibiendo la transmisión nociceptiva. Esta inhibición puede ser pre y post, y puede ser bloqueado por antagonistas serotoninérgicos, lo cual indica que dicha analgesia inducida está medidada por la liberación de serotonina.
Otra conexión descendente importante se origina también en el PGA proyecta a regiones del puente, incluyendo el locus coeruleos y la región parabraquial. Su estimulación origina proyecciones noradrenérgicas al asta dorsal de la médula espinal. Estudios experimentales han mostrado que agonistas noradrenérgicos aplicados en el asta dorsal de la médula inhiben la respuesta de neuronas nociceptivas primarias así como del tracto espinotalámico.
Hay otras sustancias muy importantes en la transmisión dolorosa que se llaman péptidos opiáceos (encefalina, dinorfina, beta-endorfina…), concretamente, las interneuronas contienen este tipo de sustancias que actúan a nivel presináptico en las vías serotoninérgicas, ya que si se administra naloxona (antagonista opiáceo) se suprime la analgesia. Por lo que respecta a las vías noradrenérgicas, no está clara que su acción se encuentre mediada por opiáceos.
Acupuntura
Se cree que parte de su acción analgésica esta mediada por las vías descendentes inhibitorias localizadas en el tallo cerebral, sustancia gris central y regiones del sistema límbico y que podrían actual sobre receptores opiáceos endógenos
Efecto placebo.
Puede venir desencadenado por la liberación de opiáceos endógenos.
Capsaicina y analgesia.
La capsaicina es una sustancia que se encuentra en las guindillas, actúa en los receptores vaniloides de las fibras nociceptivas cuando se produce alguna inflamación, la exposición prolognada de la capsaicina provoca la destrucción de los receptores así como las terminales periféricas nociceptivas.
Trastornos dolorosos.
Dolor central
Veremos tratamientos mas invasivos que se aprovechan de la anatomía de este sistema y consisten en la interrupción de las vías ascendentes de la transmisión del dolor a diferentes niveles.
-
En las raíces dorsales de la médula: bloquea toda la información, no es muy aconsejable. Se te puede ir la mano…
-
En las vías neurales donde la información dolorosa se encuentra aislada, en la zona anetrolateral. Solo es apropiado cuando el dolor se encuentra muy localizado.
-
En el tálamo, se procesan los dos tipos de dolor , se suele intervenir en el tálamo medial porque el dolor crónico está más aislado, aunque a veces no se consigue y se produce el síndrome del dolor central.
Dolor referido y dolor proyectado.
El dolor referido es que te duele una parte alejada de los tejidos que realmente producen dolor
En el dolor proyectado las fibras nerviosas encargadas de transmitir el dolor pueden no morir.
TEMA 4: NEUROBIOLOGÍA DE LA VISIÓN
Introducción: el ojo
El ojo es el encargado de focalizar las imágenes exteriores sobre una zona fotosensible denominada retina.
Parámetros físicos de la luz (el estímulos físico de la visión): Amplitud (intensidad del estímulo), Frecuencia, Longitud de onda y Saturación.
En la focalización el ojo debe acomodarse al estímulo y son importantes dos componentes:
-
la córnea: es la parte más externa del ojo, transparente y deja pasar la luz. La cantidad de luz que llega está regulada por la pupila, formada por la apertura del iris, que consiste en un aro de músculos situados detrás de la córnea.
-
el cristalino: está situado detrás del iris y consiste en una serie de capas transparentes que pueden modificar su forma gracias a la musculatura ciliar. Cuando está aplanado (distenido) enfoca objetos distantes, mientas que cuando se contrae enfoca objetos próximos. Este proceso se denomina acomodación y se lleva junto con el de convergencia (proceso que mantiene la fijación de ambos ojos sobre el mismo objetivo)
Posteriormente la luz se introduce en el humor vítreo, para terminar sobre la retina, lugar donde se localizan los fotoreceptores.
La retina: contiene varias capas (fotorreceptora, de células bipolares, Células horizontales y amacrinas, Células ganglionares )
Los fotorreceptores forman sinapsis con las células bipolares, las cuales enlazan con las ganglionares, cuyos axones llegan al cerebro. Las células horizontales y amacrinas transmiten información con el resto de capas.
La transformación será llevada a cabo por los fotorreceptores ( conos y bastones). La retina humana tiene aproximadamente 120 millones de bastones por 6 millones de conos), a pesar de esto, los conos nos proporcionan la mayor parte de información, tanto es así, que por ejemplo, la fóvea sólo dispone de conos. Hay de tres tipos: azules, verdes y rojos. En el extremo externo de cada receptor se encuentran unas moléculas llamadas fotopigmentos, que son sensibles a la longitud de onda, los bastones utilizan la rodopsina, el fotopigmento más conocido de los conos es la yodopsina.
En la oscuridad los conos y bastones se encuentran en estado de reposo, el potencial de membrana de la célula es ligeramente positivo, esto es debido a que en una situación de reposo (oscuridad), en la célula entra sodio debido a que tiene sus canales iónicos abiertos. Si en ese momento se presenta un estímulo luminoso el fotopigmento lo absorve y se subdivide en dos componentes: opsina y retinal, la opsina inicia una secuencia bioquímica que hace que se cierren los canáles iónicos. Esto produce la hiperpolarización de la célula receptora (cono o bastón ) que exceptualmente es un potencial negativo que será transmitido por potenciales graduados de unas células a otras hasta llegar a las ganglionares en dónde se producirán potenciales de acción que irán al cerebro. Cuando la luz desaparece el fotopigmento se regenera.
-
Anatomía y fisiología del sistema visual en sus diferentes niveles.
La retina puede dividirse en dos regiones, la nasal y la temporal.
El campo visual externo puede subdividirse en hemicampo derecho y hemicampo izquierdo.
La imagen del hemicampo izqueirdo se proyecta invertida a la región temporal de la retina derecha y a la región nasal de la retina izquierda, la imagen del hemicampo derecho se proyecta invertida a la región temporal de la retina izquierda y a la nasal de la derecha. La luz que se origina en la región central llega a ambos ojos en la zona binocular.
Los axones de las células ganglionares forman en nervio óptico. Sus células responden de manera diferente según se encuentran en el centro o en la periferia:
-
Las células ganglionares X o parvocelulares: Se localizan en la fóvea. Forman las tres cuartas partes de las células ganglionares de la retina. Sus cuerpos celulares son de tamaño medio y sus campos receptivos pequeños y concéntricos, tienen un centro y una periferia antagónicos. Existen dos tipos:
-
De centro on (on-off): centro excitatorio y anillo inhibitorio.
-
De centro off (off-on): centro inhibitorio y anillo excitatorio.
-
Las células ganglionares Y o magnocelulares: localizadas en la periferia de la retina. Tienen grandes campos receptivos circulares, si la luz cae de lleno en todo su campo responden ( cosa que no pasaba en las parvocelulares, porque al tener dos campos antagónicos se anulan)
Las partes temporales del nervio óptico se dirigen hacia el cerebro por el lado del ojo ipsilateral. Las nasales cruzan al lado contario. Ese cruce se produce en el quiasma óptico. A partir de aquí se forma el tracto óptico que lleva información de ambos ojos. Sus relevos son
-
A nivel del mesencéfalo, en el colículo superior (movimientos oculares y atención)
-
A nivel hipotalámico: a) hipotálamo lateral (nutrición) b) núcleo supraquiasmático (reloj interno y ritmos circadianos).
Si bien, su relevo más importante será en el tálamo, concretamente en el NGL, el NGL está formado por seis capas de estratos celulares:
-
Sistema magnocelular: Las dos capas celulares mas ventrales (1 y 2) contienen cuerpos celulares grandes, reciben aferencias de las células ganglionares tipo Y (implicadas en la visión de la forma y el movimiento)
-
Sistema parvocelular: El resto de las capas (3-6) tienen cuerpos celulares más pequeños, reciben información de las células ganglionares tipo X (codifican el color y los detalles)
Los axones de las células magnocelulares (Y) y parvocelulares (X) cel NGL establecen contacto sináptico con la corteza visual primaria o córtex estriado (área 17 o V1). La corteza estriada también está formada por 6 estratos; las células geniculadas parvocelulares y magnocelulares proyectan a la capa IV C (manteniéndose la segregación ya que:
-
las neuronas parvocelulares mandan información a la capa IV C Beta. Posteriormente proyectan a las capas II y II (blobs e interblobs)
-
las neuronas magnocelulares sinaptan con la capa IV C alfa. Posteriormente mandan información a la capa IV B.
Existen tres canales relativamente independientes de procesamiento de la información visual:
-
Movimiento (canal magnocelular): este canal comienza en las células ganglionares de tipo magnocelular de la retina y que llega a la capa IV B del córtex estriado. Las células de esta región (células simples) responden más adecuadamente a estímulos como líneas o barras y que se encuentran en una orientación espacial determinada. Si se cambia la orientación del estímulo disminuye la respuesta.
-
Forma del objeto (canal parvocelular-interblob) se inicia en las células ganglionares parvocelulares de la retina y que llega a las regiones interblob de la capas II y III del córtex estriado. Denominaron a las células de la zona intergotas como complejas porque son insensibles a las longitudes de onda y selectivas a la orientación. No existe distinción entre regiones ON y OFF.
-
Color (Canal parvolelular-blog): tiene su origen en las células ganglionares parvolelulares de la retina y que termina en los blob o gotas de las capas II yIII del córtex estriado. Estas células son sensibles a las longitudes de onda y no muestran sensibilidad a la orientación del estímulo.
-
Otras regiones corticales: En el córtex hay al menos 32 representaciones de la retina.
-
Área V2: se visualizan una especie de bandas anchas, estrechas y pálidas.
-
Las bandas anchas reciben aferencias de la capa IV B (Canal Magnocelular) proyecta a V3 y a V5 o MT . V3 se encarga de los aspectos relacionados con la orientación o la visión de la forma en blanco y negro, V5 está especializada en la visión del movimiento.
-
Las bandas estrechas reciben información de los blobs (Canal Parvocelular-Blobs) mandan información a una región llamada V4 o giro fusiforme
-
La región pálida recibe señales de las regiones interblobs (Canal Parvocelular- Interblobs) también manda información a V4.
A partir de aquí se van a formar dos corrientes o sistema de procesamiento,
-
Uno ventral(¿Qué es el objeto?): terminando en la corteza del lóbulo temporal inferior, está representada por el sistema parvocelular, donde existen dos relevos: TE y TEO. La zona TEO recibe su información en buena parte de V4 y no se encarga de la percepción del color sino del reconocimiento de los objetos. La zona TE utiliza esa información para los sentimientos y las emociones.
-
El otro es ascendentey dorsal(¿Dónde se localiza?), y termina en la corteza del lóbulo parietal posterior. Se transmiten a través de las capas magnocelulares. Se encarga de aspectos como el movimiento y la organización tridimientosional. Esta información nace de las células grandes retinianas tipo M a V1, estas señales se elaboran en el área temporal medial (TM o V5) y la región temporal superior medial (TSM)
Visión del color
Thomas Young señaló que el color está mediado por tres clases de fotorreceptores, los fotopigmentos son sensibles a longitudes de onda cortas, para ver el azul, a longitudes de onda medias, para ver el verde y a longitudes de onda largas para ver el rojo; estas pruebas confirmaron que la visión de los colores se inicia a nivelde los fotorreceptores de la retina.
pero no a los colores como proponía la teoría tricromática. Otro problema es que no pude responder a la cuestión de que algunos colores parecen mezclarse y otros en los que parece imposible.
Estas dificultades llevaron a Hering en 1877 a proponer la teoría de los procesos oponentes. Dicha teoría se va a centrar en la células ganglionares de la retina, se han identificado dos tipos de células (rojo-verde/azul-amarillo). También existiría un tercer tipo de célula (blanco-negro)
Para la percepción del color dependemos inicialmente de la longitud de onda que reflejan las superficies de los objetos, a partir de ellas deducimos los colores. Esa longitud de onda no solo viene determinada por el color del objeto sino también por las diferentes longitudes de onda de la luz ambiental que pueden variar en función del tiempo atmosférico y lugar (constancia del color).
Eso hizo que Land propusiese la teoría de la constancia del color o teoría retinex, la cual señala que la visión del color nos e deduce exclusivamente de lo que refleja una superficie sino que el sistema nervioso es activo, crea el color y para deducir el color debe comparar una superficie con lo que reflejan las superficies del entorno. La zona aparentemente implicada fue V5, si se destruye esa región se dejan de ver los colores.
Efectos visuales de las alteraciones de las vías y centros sensoriales.
-
La lesión que afecta a la retina temporal del ojo derecho. La persona no ve la parte central del hemicampo izquierdo.
-
Lesión que afecta a la retina nasal del ojo izquierdo. La persona no ve la parte lateral del hemicampo izquierdo.
-
Si la lesión destruye el nervio óptico derecho, se pierde visión de la parte central del hemicampo izquierdo así como la parte lateral del hemicampo derecho (la visión del ojo derecho)
-
Si la lesión destruye el quiasma óptico se pierde la visión de la parte lateral del hemicampo derecho.
-
Si la lesión destruye el tracto óptico derecho, se pierde la visión del hemicampo izquierdo, o lo que es lo mismo, de la visión derecha de cada retina. Lo mismo sucede si la lesión se produce en los núcleos geniculdos del tálamo
-
Si la lesión destruye la región V1 se produce visión ciega.
Zonas cerebrales relacionadas con la percepción visual. Las agnosias.
-
acromatopsia, que es la incapacidad para reconocer colores. La zona afectada sería V4
-
acinetopsia, las personas no pueden ver el movimiento, y la zona implicada es V5
-
prosopagnosia es una agnosia visual que imposibilita el reconocimiento de caras. Se han identificado v arias células en el lóbulo temporal inferior.
TEMA 5: LA AUDICIÓN
Parámetros de la audición.
La frecuencia del sonido se define como el número de moléculas de aire que pasan por nuestros oídos en cada segundo. La frecuencia se percibe como tono, así variaciones en la frecuencia de un sonido producen variaciones en la percepción del tono.
La intensidad lo determina lo alto o fuerte que percibimos
El timbre proporciona información sobre la naturaleza de un sonido en particular.
La máxima sensibilidad auditiva se da en la franja que va desde los 1000 Hz a los 4000 Hz, rango donde percibimos el lenguaje. Esa sensibilidad a las diferentes frecuencias se mide con los audiogramas.
Estructura del sistema auditivo del oído
-
Anatomía
El oído puede ser subdividido en:
-
oído externo: pabellón y canal autidivo
-
oído medio: membrana timpánica y osículos
-
oído interno: ventana oval y cóclea (órgano de Corti)
La parte visible del oído consiste en una zona cartalaginosa recubierta de piel, denominada pabellón auditivo, cuando la onda sonora lo traspasa, llega al canal auditivo para pasar a la membrana timpánica; donde hay conectados una serie de huesecillos denominados osículos que forman el oído medio:
-
El martillo se encuentra conectado a la membrana timpánica, al mismo tiempo que forma una conexión rígida con el yunque, este, a su vez mantiene una conexión flexible con el estribo. La base del estribo se mueve hacia adentro y afuera sobre la ventana oval, transmitiendo las vibraciones del sonido a los fluidos de la cóclea dentro del oído interno. La principal función de los huesecillos es la amplificación del sonido.
La vibración de los huesecillos se encuentra limitada por dos estructuras musculares que se encargan de proteger a las células receptoras del oído interno de los sonidos fuertes: uno de esos músculos (el tensor del tímpano) se encuentra adosado al sistema martillo y es controlado por el trigémino. La otra estructura muscular (el estapedio) esta gobernado por el VII par craneal (facial) y se encuentra adosado al estribo.
Cuando se desde el sistema osicular se transmiten sonidos fuertes hasta el sistema nervioso central, tras un breve periodo se produce un reflejo que provoca la contracción del músculo estapedio y tensor del tímpano. El tensor del tímpano tracciona hacia adentro el mango del matillo mientras que el estapedio lo hace hacia fuera. Estas dos fuerzas que se oponen entre sí, hacen que el sistema osicular desarrolle un alto grado de rigidez lo que reduce la conducción osicular del sonido.
La cóclea tiene forma espiral, en la base hay dos agujeros cubiertos de membrana: la ventana oval y la ventana redona. Si se corta la cóclea transversalmente, se puede ver que se divide en tres canales : vestibular, media y timpánica. La membrana de Reissner separa la escala vestibular de la escala media, la membrana basilar separa la escala timpánica de la escala media.
Sobre la membrana basilar se encuentra el órgano de Corti, que contiene las neuronas receptoras autitivas. Por encima del aparato de Corti se encuentra la membrana tectorial.
El caracol se encuentra repleto de fluido, el fluido de la escala vestibular y timpánica se encuentra en contacto un agujero denominado helicotrema. El fluido se llama perilinfa. la escala media se encuentra incomunicada y repleta de fluido que recibe el nombre de endolinfa.
Cuando el sonido entra por el pabellón autidivo, estos mueven a la ventana oval. El movimiento hacia adentro de la ventana oval empuja la perilinfa de la escala vestivular, esto hace que dichos movimientos se transmitan a la ventana redonda que se mueve hacia adentro y afuera de forma opuesta a los movimientos de la ventana oval. Es decir, cuando la base del estribo empuja hacia adentro, la membrana de la ventana redonda se dobla hacia fuera.
El órgano de Corti está constituido por una serie de células receptoras que están ancladas a la membrana basilar, y que se diferencian en dos tipos: externas e internas. Las células ciliadas internas son las más imortantes en la audición, ya que proporcionan el 95% de las fibras aferentes auditivas. Sus cilios (pelos) no tocan a la membrana tectorial, al contrario de lo que ocurre con las células ciliadas externas.
Las células ciliadas se encuentran conectados a través de fibras neurales (cuyos cuerpos celulares están en el ganglio espiral) sus axones forman el nervio auditivo (VIII par craneal).
-
Fisiología
Las células ciliadas internas se mueven por el desplazamiento del líquido de la escala media. Ese movimiento hace que se produzca una apertura de los canáles iónicos en sus terminales. Los cilios se abren como compuestas y se produce una entrada de iones que abundan en la endolifa (K ). Su entrada produce una despolarización que activa los canales de Calcio. La entrada de Calcio desencadena la liberación de un neurotransmisor denominado Glutamato, que activará a las fibras del ganglio espiral que forman el nervio auditivo.
Investigaciones recientes sugieren que las células ciliadas externas juegan un papel fundamental en la transducción del sonido, pues modifican las relaciones físicas entre las membranas cocleares cambiando la longitud de las proteínas motoras de las células, de esta manera la membrana basilar se separa de la lámina reticular y la membrana tectorial. Ruggero y cols. Administraron Furosemida en animales experimentales, esta sustancia decrece temporalmente la transducción que se produce por el desplazamiento de los cilios y se piensa que es por la inactivación de las proteínas motoras de las células ciliadas externas. Otra evidencia es que ciertos antibióticos pueden llevar a la sordera y parece deberse a la destrucción selecctiva de las células ciliadas externas.
-
Neuroanatomia del sistema autidivo
El VIII par craneal tiene su primer relevo a nivel del tallo cerebral, en los núcleos cocleares. Estas células envían proyecciones a la oliva superior, este núcleo recibe información de ambos núcleos cocleares, se produce una interacción binaural. Desde la oliva superior mandan información al colículos inferior del mesencéfalo. Esta región a su vez envía axones al núcleo geniculado medial del tálamo, el cual proyecta al córtex autivo, localizado en la circunvolución superior del lóbulo temporal.
Percepción de la frecuencia del sonido: organización tonotópica de las células auditivas y frecuencia óptima.
-
Teoría temporal o de la frecuencia (Tª de Volley o de la descarga)
Enfatiza las relaciones entre la frecuencia del estímulo y el cuadro de disparo de la célula, de acuerdo con esto la neurona modificará o alterará su tasa de disparo ante el cambio en la frecuencia del estímulo.
Se ha demostrado que esta idea es válida ante frecuencias bajas, pero el sistema nervioso capta frecuencias de 15000 ciclos/seg y no hay ninguna célula que llegue a tal tasa de disparo.
-
Teoría de Campo o Teoría Espacial (Tª de Von Békesy)
Señaló que todas las frecuencias se repartirán a lo largo del caracol, de modo que las células basales se encargan de las frecuencias altas y las más profundas de las bajas.
Cuando se presenta un estímulo se va a producir una acción sobre la membrana basilar diferencie en cada lugar según la frecuencia del sonido. Si la frecuencia es alta, la base de la membrana basilar vibrará en buena medida proporcionando la mayor parte de la energía, y la onda no se propagará más lejos. Sin embargo, los sonidos de baja frecuencia generará ondas que viajan a lo largo de la membrana, proporcionando la mayor parte de la energía en su extremo, en definitiva, el sistema nervioso sabrá qué frecuencia ha llegado en función de la célula que se ha activado. Existen varias pruebas que apoyan esta idea.
Algunos de los enfermos de tuberculosis que tomaban antibióticos en grandes cantidades se dañaban las células ciliares, esta destrucción siempre comenzaba por la base de la cóclea y era progresiva, y dichos individuos percían ciertas frecuencias y no otras.
Otra prueba proviene de los estudios del funcionamiento de las células auditivas. Se observó que cada célula tenía una sensibilidad a las frecuencias en forma de uve(V)
Otra prueba es la organización tonotópica, fibras del nervio autidivo que conectan con células sensoriales cercanas al extremo de la membrana basal, tienen características de baja frecuencia, las que se encuentran conectadas con células sensoriales cerca de la base de la membrana basal poseen características de alta frecuencia. Estos mapas se siguen produciendo en los siguientes relevos: núcleo geniculado medial y córtex auditivo.
Esta teoría tiene dificultades a la hora de explicar la percepción de frecuencias bajas (200Hz) ante estos casos sí parecen existir células individuales que pueden seguir ese ritmo de disparo. Según la teoría de la frecuencia.
La combinación de ambas teorías se denomina teoría dúplex.
-
la intensidad del estímulo.
Este proceso va a depender de que las células que procesan las frecuencias lo hagan en función de la teoría espacial o temporal.
Si la frecuencia es alta, se activará la célula correspondiente de la base de la membrana basilar que dispara a un ritmo determinado. Si la intensidad es mayor, disparará más rápido.
Cuando la frecuencia es baja, las células la procesan disparando al ritmo de la frecuencia del estímulo, no puede utilizar su tasa de disparo para procesar incrementos en intensidad porque esto lo utiliza para procesar la frecuencia. Ante intensidades elevadas, mayor número de células implicadas.
-
Mecanismos de localización auditiva del sonido.
Las pistas para la localización auditiva son las diferencias entre los oídos en cuanto a la intensidad del sonido y el tiempo de llegada:
Un sonido que no procede ni de delante ni de detrás de la persona llegará a un oído antes que a otro, esta disparidad se denomina Delta “t”.
Los sonidos que no provengan ni de delante ni de detrás llegarán a los oídos con distinta intensidad. Esta disparidad de intensidad se denomina Delta “fi”
La utilización de ambas no es indiscriminada, sino que la mayoría de las especies suelen tomar una u otra, debido a las características de la cabeza, la disparidad temporal tiene una validez menor cuanto menor sea la cabeza de esa especie. Si muy es muy pequeña la posibilidad de disparidad temporal es nula. Y utilizan como índice la disparidad de intensidad (el efecto pantalla de su cabeza) esto solo es posible con frecuencias altas, por eso no son muy sensible a las bajas. El mecanismo clave de todo esto es el complejo Olivar Superior (si se destruye se elimina la capacidad de difeernciar la localización de los sonidos). Esta estructura se subdivide en dos subnúcleos: lateral y medial. Cada uno se dedica a las diferentes especialidades. Si se usa disparidad temporal el núcleo medial estará mas desarrollado y si se usa la disparidad de intensidad el núcleo lateral estará mas desarrollado.
Por su parte, la corteza auditiva es esencial en la tarea de localización, organiza el espacio auditivo para actuar en el.
-
Repercusiones comportamentales del daño en el sistema sensorial auditivo.
La sordera se produce generalmente por problemas en o cerca de la cóclea. Las repercusiones son de dos tipos: sensoriales y perceptivas.
-
Sensoriales:
Sordera de conducción: se produce cuando las alteraciones auditivas están asociadas con patologías de las cavidades auditivas externa o media. La pérdida de audición se basa en la incapacidad para excitar mecánicamente la cóclea. El oído medio está conectado por la trompa de Eustaquio con el tracto respiratorio superior, por lo que infecciones de garganta pueden acceder al oído medio. Todas estas anomalías de conducción pueden ser tratadas.
Sordera nerviosa: asociada con la pérdida de neuronas en el nervio auditivo o en las células ciliadas de la cóclea. Las causas son bastante variadas ( desde exposición a mucho ruido, tumores, antibióticos…)Si la cóclea o el nervio auditivo de un oído son completamente destruidos la sordera de este oído es absoluta. Sin embargo una pérdida parcial es más común, en estos casos se utiliza un amplificador del sonido.
Si hay lesión a partir del núcleo olivar, no se produce sordera, puesto que la información auditiva es bilateral, a nivel del tálamo (cuerpo geniculado medial ) lo mismo, y algo parecido ocurre en la corteza, pues se produce una sordera, esta es recuperable.
Hay un trastorno conocido con el nombre de Tinnitus, en los que una persona escucha sonidos en los oídos en ausencia de algún estímulo sonoro. No se sabe la causa (puede ser por una vibración constante de los huesos del oódo medio debido a algún daño de los músculos que los controlan) ni la solución ( a día de hoy solo se puede “enmascarar” el sonido)
-
Perceptivas
Se conocen como agnosias auditivas.
Agnosias como sonidos no verbales: no pueden dar un significado a lo que oyen. Se producen por lesiones en la corteza (en el lóbulo temporal medial y superior del hemisferio derecho, generalmente son bilaterales)
Agnosias musicales: No aprecian las características de la música, también por daños temporales mediales. O no reconocen melodías familiares, por daños localizados en el hemisferio derecho
Agnosias a sonidos verbales: no se puede comprender el lenguaje, pero pueden hablar y escribir correctamente. Se deben a lesiones vasculares que afectan bilateralmente a los lóbulos temporales.
TEMA 7: FACTORES PERIFÉRICOS EN EL CONTROL DE LA NUTRICIÓN
Introducción
Desde la perspectiva biológica se considera que la conducta nutritiva es una conducta motivada, dirigida a conseguir la energía necesaria para mantener el equilibrio homeostático del organismo. Se asume que el organismo debe contar con mecanismos que le permitan detectar señales indicadoras de la existencia de un déficit y desencadenar una serie de comportamientos dirigidos al restablecimiento del equilibrio nutritivo. También ha de ser capaz de captar las señales que le informan que el déficit ha sido corregido.
Factores implicados en el inicio de la ingesta.
-
Teoría glucostática:
Carlson en 1916 sugirió que la señal fisiológica que determina el inicio de la ingesta está constituida por la cantidad de glucosa presente en el sistema circulatorio, en los casos de diabetes, un síndrome caracterizado por la hiperglucemia dicha sensación persiste.
En 1955 Mayer midificó la idea avanzada por Carlson y presentó la teoría glucostática según la cual el mecanismo regulador crucial no consistía en la concentración de glucosa sino en la tasa de utilización de la misma.
-
Teoría lipostática
Propuesta por Kennedy en 1953 defiende que las reservas en las grasas pueden determinar e influir en la nutrición, manipulaciones farmacológicas que bloquean la utilización de los ácidos grasos (mercaptoeacetato o metilpalmoxirato, estimulan la ingesta de alimentos de forma efectiva.
-
Teoría aminostática
Mellinkoff plantea que un exceso o déficit en los niveles de animoácidos en el plasma son fenómenos que afectan al inicio o inhibición de la ingesta respectivamente.
Estos tres grupos de macronutrientes pueden contribuir a la hora de general la sensación de déficit, sin embargo, deben existir sistemas biológicos que detecten en cada momento la disponibilidad de dichos nutrientes. Los datos actuales demuestran que existen receptores implicados en la detección de los déficits energéticos ubicados tanto a nivel central como periférico. Los mecanismos que controlan la disponibilidad de glucosa residen en el encéfalo, los receptores sensibles a déficits lipídicos o carencias de otros carbohidratos parecen localizarse a nivel hepático y posiblemente intestinal. En este último caso la información es remitida a través de aferencias en el nervio vago.
Parece que en el inicio de la ingesta también están implicados factores de naturaleza endocrina. La grelina es una hormona peptídica, es un potente estimulador del apetito en animales cuando se administra exógenamente. En condiciones de ingesta espontánea la hormona se libera desde la mucosa gástrica, unos 20 o 30 minutos antes de empezar a comer, otra evidencia es que los pacientes operados con un bypass las concentraciones de la hormona disminuyen, por lo tanto comen menos que otros pacientes que han sido tratados con otros métodos quirúrgicos.
Factores periféricos en el cese de la ingesta
Hay que diferenciar entre dos procesos:
-
Saciación o saciedad a corto plazo es el mecanismo responsable de que un animal deje de tomar alimento durante un episodio de ingesta
-
Saciedad o saciedad a largo plazo hace referencia a la ausencia de déficit hasta el inicio de una nueva comida.
La naturaleza de las señales implicadas en estos procesos es diferente pues los procesos que indican la finalización de la ingesta, deben proceder de los niveles del tracto digestivo, que serían comunicados al sistema nervioso central a través del nervio vago, por el contrario, las señales que regulan la saciedad están más relacionadas con efectos metabólicos postabsortivos como la disponibilidad de nutrientes en sangre
-
Factores orosensoriales.
Han sido estudiados en con una técnica denominada “alimentación ficticia” consiste en impedir que el alimento alcance el estómago, por lo que es drenado inmediatamente al exterior. En estas circunstancias los sujetos no manifiestan la típica saciedad pues el tamaño de la comida es mayor y el tiempo transcurrido entre dos comidas mucho menor.
A pesar de ello, la experiencia oral puede ser importante, esta afirmación se basa en un fenómeno conocido como “saciedad sensorialmente específica” que consiste en una temprana reducción de la ingesta cuando el alimento ofrecido es siempre el mismo. En cambio, cuando la dieta es variada, la ingesta aumenta y el cese aparece mucho mas tarde.
-
Factores gástricos.
La distensión del estómago es uno de los mecanismos responsables en la finalización de la ingesta, en estudios experimentales que permiten distendir la cavidad gástrica durante un episodio de ingesta así lo confirman. La gastroplagia, por ejemplo, es una técnica que reduce el tamaño del estómago; algunos sujetos terminan incrementando la frecuencia de las comidas. Las señales gástricas son conducidas a través del nervio vago.
-
Factores intestinales
La participación de factores intestinales en el cese de la ingesta es defendida en la actualidad por muchos científicos. Algunos estudios han demostrado la contriución postgástrica a la saciedad a corto plazo administrando nutrientes directamente en el duodeno u otros segmentos del intestino en una situación de alimentación ficticia, estos estudios han sido criticados por que la finalización de la ingesta parece corresponder mas a factores de tipo aversivo que a efectos saciadores.
Otros autores defienden que las señales de saciación podrían estar mediadas por la liberación de ciertos péptidos gastrointestinales durante el paso del alimento. Muchos de estos péptidos, además, tienen receptores en fibras nerviosas perfiéricas y/o en regiones del SNC implicas en la regulación de la ingesta.
La primera hormona propuesta fue la colecistoquinina (CCK) un polipéptido que se libera al torrente sanguíneo desde la mucosa duodenal cuando los nutrientes entran en este segmento del tubo digestivo.
Algunos estudios, sin embargo, ponen esta hipótesis en tela de juicio, existen trabajos que demuestran que las dosis de CCK que inhiben la ingesta son superiores a las dosis fisiológicas que se liberan pos-prandialmente.
Otra hormona candidata es el glucagón, el péptido liberador de gastrina (grp)/bombesina, el péptido relacionado con el gen de la calcitonina (CGRP) o la amilina. Todos reducen la ingesta cuando se administran exógenamente.
Recientemente ha sido implicado el péptido YY (PYY) se libera desde el intestino delgado distal y el colo en en proporción a las calorías ingeridas, sus niveles aumentan durante las comidas. La administración exógena de PYY reduce la ingesta de alimento en animales y humanos.
-
Factores hepáticos
La última etapa ocurre en el hígado, que es el primer lugar donde fluyen la mayoría de los nutrientes a medida que se va produciendo la absorción desde el tubo digestivo.
Las señales de saciación generadas en los distintos niveles del tracto gastrointestinal son comunicadas al SNC a través de aferencias contenidas en el nervio vago. Este nervio proyecta casi exclusivamente al núcleo del tracto solitario (NTS) un núcleo troncoencefálico que, comunica directa e indirectaemnte con estructuras troncoencefálicas, diencefálicas y telencefálicas implicadas en el control de la ignesta de alimento.
Factores periféricos en la regulación del peso corporal.
Existe un acuerdo entre los autores en que la teoría lipostática ofrece un modelo más adecuado para la regulación de la homeostasis energética. Según esta teoría, el cerebro en condiciones de normalidad, controla constantemente la cantidad de energía almacenada en el cuerpo y actúa para impedir cambios en la misma. Para ello se sirve de señales periféricas que le informan del estado de las reservas.
La señal responsable no se conoce con exactitud. Algunos investigadores plantearon la posibilidad de que dichas señales puedan ser indirecta (el cerebro mediría las reservas en función de los macronutrientes que hay en sangre) otra posibilidad (la directa) supone que el cerebro es informado a través de productos intermediarios que se liberan desde la periferia en proporción a la grasa almacenada.
La hipótesis anterior no es actual. Herbey lo estudió en 1959 con animales parabióticos (parejas de sujetos unidos por la musculatura y la piel de los costados) de manera que se desarrollan conexiones en el sistema vascular, pero no conexiones nerviosas. Pudo comprobar que cuando los dos sujetos eran normales o obesos, cada animal mantenía el nivel de ingesta usual y el peso corporal no se modificaba en ninguno. Sin embargo si uno era obeso y el otro normal, este último se volvía hipofágico y perdía peso. Hervey lo interpretó como que algún factor humoral proporcional a la obesidad era transferido del animal grueso al delgado alcanzando las áreas del cerebro que controlan la ingesta de alimento. En el obeso, este factor no tendría efecto, dado que estas áreas o parte de ellas estarían lesionadas. En los animales normales, el cerebro respondería como si el animal tuviera más grasa de la que en realidad posee induciendo hipofagia y pérdida de peso.
Douglas estudió los datos obtenidos por Hervey con ratones genéticamente obesos y fenotípicamente iguales (ob/ob y db/db). Demostró que:
-
Si uno de los sujetos de la pareja era db/db y el otro normal, este último dejaba de comer y perdía peso.
-
Si por el contrario el normal se unía con un ratón ob/ob era este último el que adelgazaba aunque sin dejar de ser obeso.
-
Si los animales conectados eran ob/ob y db/db, el ob/ob dejaba de comer y perdia peso
En base a esto Coleman propuso la existencia de un factor saciador humoral (leptina)que estaba ausente en ratones ob/ob y que los db/db producían en exceso aunque no eran sensibles a él. Sin embargo, la ingesta no estimula la leptina; por lo tanto se ha propuesto que esta hormona podría liberarse de manera proporcional a la cantidad de grasa corporal.
Por otra parte, se ha demostrado que la leptina alcanza el cerebro a través de mecanismos de transporte específicos y saturables de forma que la cantidad del péptido en el SNC es proporcional a la concentración detectada en el plasma.
Muchas de las características de la leptina son compartidas por la insulina, una hormona, de origen periférico (hepático) se ha propuesto que tanto la leptina, como la insulina podrían ser señales que informan al cerebro sobre el estado de las reservas de grasa.
El conocimiento de la leptina hizo albergar esperanzas en personas con trastornos nutritivos. Pero la obesisdad humana no se puede atribuir a un solo factor, pues dichos sujetos poseen niveles elevados de esta hormona en sangre, lo que indica que continúa produciéndose adecuadamente en proporción a la cantidad de grasa, el problema podría estar en su transportador, pues se ha demostrado que la proporción de leptina en sangre y la proporción de leptina en el LCR que existe en sujetos con peso normal no se observa en pacientes obesos, siendo los niveles intracerebrales anormalmente bajos.
Existen dos tipos de factores endocrinos periféricos implicados en la regulación de la conducta nutritiva. Por un lado están los reguladores a corto plazo (CCK) que afectan al transcurso de una comida, por el tro están los reguladores a largo plazo, como la leptina o la insulina, cuya secreción se asocia con cambios metabólicos.
Interacción de los sistemas implicados en la regulación de la ingesta y peso corporal
La homeostasis energética está regulada por un cambio sistema de estructuras interconectadas que se localizan a distintos niveles del sistema nervioso. En estas estructuras pueden direferenciarse:
-
circuitos anabólicos (relacionados con el inicio de la ingesta)
Entre los sistemas que forman parte de este circuito destacan los integrados por el sistema POMP/CART que liberan dos neuropéptidos anorexigénicos, el alfa-MSH y el CART.
-
circuitos cababólicos (relacionados con la inhibición en la ingesta)
Destacan las vías integradas por el neuropéptido Y (NPY) y la proteína relacionada con el agouti (AgRP).
Dichos circuitos se encuentran en el núcleo arqueado del hipotálamo, un centro libre de barrera y por lo tanto accesible a las sustancias que están en la sangre.
En estudios recientes se ha demostrado que las neuronas que coexpresan NPY/AgRP como las neuronas POMP/CART del núcleo arqueado poseen receptores para la leptina y la insulina, y su actividad está regulada por estas hormonas, aunque de manera opuesta. Las nueronas que expresan NPY y AgRP son inbhidias por la leptina/insulina (se activan cuando los niveles de grasa disminuyen) las del sistema POMP son activadas por ambas hormonas (se inhiben cuando las reservas de grasa disminuyen y los niveles de leptina e insulina en sangre descienden)
TEMA 8: TRASTORNOS COMPORTAMENTALES HIDROMINERALES: EL CASO DE LA DIABETES INSÍPIDA CENTRAL.
Distribución del agua entre los diferentes compartimentos celulares y su regulación con la sed
Por término medio, una persona sana pierde una media de 1,5 litros al día, este déficit será compensado por la toma de agua, comida o el metabolismo de la dieta.
Dos terceras partes del agua corporal se encuentra en el interior de las células en condiciones estables, el tercio restante en compartimentos extracelulares.
El soluto aparece en distintas concentraciones según se localice en el interior o en el exterior de la célula, así el potasio es muy abundante en el interior (141 mosmol/l frente a 5 mosmol/l fuera) lo mismo sucede con el sodio pero en sentido contrario (10 mosmol/l en el interior y 144 mosmol/l en el exterior)
Cualquier alteración hace que el agua y fluya de un lado hacia el oro de la membrana basándose en principios osmóticos.
Esta dinámica interna puede verse afectada por varios factores, entre ellos:
-
Hipertonicidad: que es el aumento en la concentración del medio extracelular, la célula queda deshidratada por lo que se induce a la toma de agua.
La privación de agua, consumo de alimentos ricos en sodio, o sustancias hipertónicasa como el ClNa o la sacarosa son factores desencadenantes de la puesta en marcha de mecanismos de conservación y regulación de fluidos renales y electrolitos, así como del consumo de agua originado por variables osmóticas
-
Hipovolemia: ante una reducción del líquido intravascular se desencadenan una serie de reacciones neuroendocrinas compensatorias que posibilitan la conservación y retención de fluidos corporales (respuesta antidiurética, antinatriurética y mantenimiento de la presión sanguínea en condiciones óptimas)
Una situación de hipovolemia también genera la búsqueda e ingesta de agua y sal para restaurar el déficit producido.
La sed volémica puede sr desencadenada administrando un coloide.
Cualquier alteración provoca dos respuestas adaptativas y regulatorias: el control fisiológico del medio interno y la búsqueda y consumo de agua y sal
Anatomofisiología de la sed.
Regulación osmótica de la sed: deshidratación celular
Localización de los osmorreceptores.
Verney se dio cuenta de que la administración de ClNa hipertónico en la arteria carótida interna es más efectivo que cuando eran administrados en la vena periférica. Dicha arteria solo irriga regiones prosencefálicas, lo que hizo pensar que las células sensibles a la tonicidad del plasma se encontraban ubicadas en la parte anterior del hipotálamo.
Anderson logró estimular la sed administrando ClNa hipertónico en el hipotálamo rostromedial, consiguó provocar la toma de agua en animales, si se generaban lesiones electrolíticas, los animales se volvían adípsicos.
Estudios electrofisiológicos mostraron que neuronas individuales del área preóptica lateral se activan cuando los animales son privados de agua, aunque no se puede descartar que receptores adicionales localizados en otras regiones cerebrales puedan ser los responsables de esta conducta regulatoria. En este sentido, unas pruebas experimentales mostraron que células neurosecretoras magnocelulares de los núcleos supraóptico (SON) y paraventricular (PVN) exhibían unos cambios tras la administración de ClNa hipertónico.
Sin embargo, para detectar la osmolaridad, se debe de tener acceso a la circulación sanguínea, y ninguna de estas estructuras está fuera de la barrera.
Los primeros datos destacaron el papel de la región hipotalámica situada inmediatamente rostral al extremo anteroventral del III ventrículo, la zona AV3V, pues lesiones en esa zona provocaban adipsia temporal.
Investigaciones posteriores focalizaron su atención en el órgano vasculoso de la lámina terminalis (OLVT) y el órgano subfornical (SFO). Lesiones en ambas zonas redujeron el consumo de agua.
Estudios anatómicos y eletrofisiológicos revelaron la presencia de conexiones excitatorias entre el OVLT y SFO hacia las neuronas magnocelulares hipotalámicas del SON y PVN.(las vías mas abundantes parecen estar en OVLT)
Estas zonas parecen las perfectas candidatas para detectar los cambios producidos en la tonicidad de los fluidos, sin embargo, como pasa siempre, hay otras zonas.
En este caso se puso especial énfasis en las zonas cercanas a la región anteroventral del III ventrículo, en concreto del núcleo preóptico mediano, que como se demostró recibe aferencias de OVLY y SFO.
Se podría proponer que las regiones del hipotálamo anterior detectan los cambios en la osmolaridad plasmática, para organizar sus respuestas a través del complejo SON/PVN.
Regulación volémica de la sed: deshidratación extracelular.
Las hemorragias son la causa más poderosa de deshidratación extracelular, por lo tanto de sed volémica. Debido a que la hipovolemia produce también una pérdida de sodio, hace que se pongan en marcha los mecanismos endocrinos implicados en la conservación de este mineral.
Receptores implicados en la sed volémica
James Fitzsimosn provocó hipovolemia en ratas adminstrando un coloide o desarrollando hemorragias, observó que tomaban más agua en las 6 horas siguientes que los animales controles, para evitar posibles explicaciones alternativas, como que la toma de agua es secundaria a un cuadro de poliura, les nefroctomizó, a pesar de ello observó los mismos resultados.
Las aurículas del corazón y las venas tienen neuronas sensoriales (barrorreceptores vasculares) capaces de detectar cambios en la presión sanguínea (receptores cardiopulmonares)
En sus investigaciones James, colocó un pequeño globo en la vena cava, cuando este se inflaba el flujo de sangre disminuía, lo que además de provocar sed al cabo de 30 minutos, también producía un descenso en el flujo de orina (mayor retención de agua)
Estudios neurofisiológicos han confirmado que la información proveniente de los receptores cardiopulmonares alcanza el sistema nervioso central a través de un primer relevo sináptico en el núcleo del tracto solitario (NTS) así como en el área postrema. Esta información asciende posteriormente hacia el núcleo parabraquial lateral (PBN) donde establece un segundo relevo. Desde estas regiones, la información podría modular la actividad de centros localizados en la parte anterior del III ventrículo (AV3V) así como del complejo supraóptico y paraventricular.
Sistema renina-angiotensina: sed hipovolémica y apetito por sal
Los riñones contienen células capaces de detectar la hipovolemia, su presencia hace que el riñón secrete una enzima importante, la renina, que en la sangre cataliza la conversión de proteínas en angiotensina( AII), que es la sustancia realmente crucial. Es una señal humoral de carácter dipsogénico que iba acompañada de una clara reacción vasoconstrictora periférica, así como de la liberación y secreción de determinadas hormonas.
La administración de AII provocaba una respuesta dipsogénica más eficaz y rápida que la infusión sistémica de la hormona. Ello sugería que la angiotensina debía alcanzar el cerebro y producir allí su efecto dipsogénico. Pero la angiotensina no atraviesa la barrera.
Varios científicos encabezados por Frederick Mendelsohn observaron que las neuronas del órgano subfornical (SFO) tienen receptores para la angiotensina. Pues si administración en dosis bajas provoca ingesta de agua y ClNa, mientra que la administración de saralasina (que bloquea el receptor de AII) inhibe dicha respuesta.
Una serie de estudios posteriores han demostrado que el núcleo preóptico mediano (MnPO) es muy sensible a la angiotensina, sin embargo, la angiotensina no puede acceder directamente.
La explicación es la siguiente: las neuronas del SFO serían activadas por la AII circulante y estas células enviarían señales eferentes a las neuronas del núcleo preóptico mediano utilizando también angiotensina como sustancia neurontransmisora. Entre las pruebas que avalan esta idea se encuentra que las lesiones electrolíticas del núcleo preóptico mediano interrumpían la conducta de ingesta después de la administración de AII.
Alteraciones en la ingesta de líquidos.
Los desequilibrios hidrominerales de los compartimentos intra y extracelular son capaces de generar una variedad de señales encaminadas a la ingesta de agua.
La hormona antidiurética(vasopresina) es uno de los principales mecanismos con los que cuenta nuestro organismo para poder controlar los cambios en la osmolaridad y mantenimiento del volumen sanguíneo. La vasopresina es sintetizada en los cuerpos celulares de los núcleos supraóptico y paraventricular del hipotálamo. Los axones de estas neuronas transcurren a lo largo del infundíbulo hasta llegar a la neurohipófisis el lugar dónde se va a almacenar la hormona para una posterior liberación. Suele ser liberada en situaciones de desequilibrio osmótico con el objetivo de llevar a cabo las respuestas correctoras pertinentes. Esta ejercerá su acción en los túbulos renales, promoviendo la retención de agua (respuesta antidiurética) excretándose consiguientemente una orina más concentrada.
Polidipsia secundaria: diabetes insípida.
La pérdida desproporcionada de agua se produce a causa de la insuficiente liberación de ADH (diabetes insípida central o neurogénica) o por una incapacidad renal para responder a la hormona antidiurética disponible (diabetes insípida nefrogénica).
-
Diabetes insípida central
Reducción significativa en la síntesis, almacenamiento y liberación de las hormonas neurohipofisarias (vasopresina, ADH y oxitocina, OX) lo que desencadena en una excesiva ingesta de agua.
Uno de los procedimientos que más se ha utilizado para su diagnóstico es el test de privación de líquidos. Durante su realización se observa el grado de concentración en la orina que presentan los pacientes y en el caso de la diabetes insípida se caracteriza por la osmolaridad urinaria disminuida.
Los pacientes que padecen esta enfermedad presentan una reducción notable en la intensidad de la señal emitida por las imágenes, confirman la alteración del sistema neurohipofisario.
En los últimos años se ha experimentado un gran avance en el tratamiento farmacológico con el descubrimiento de dDAVP que es un análogo de la hormona antidiurética
-
Diabetes insípida negrofrénica
Es producida por una alteración de las células del túbulo colector, lo que provocará una origna persistentemente hipotónica.
Los mejores resultados se han obtenido con la utilización de diuréticos y la restricción de sal en la dieta.
Polidipsias primarias.
Las personas consumen agua ininterrumpidamente a pesar de disponer de unos niveles de hidratación normales.
-
Polidipsia primaria dipsogénica.
El paciente informa de una sensación anormal de sed, la ingesta es regulada por factores osmóticos, pero no puede ser suprimida ya que los altos niveles de hidratación que manifiestan estos sujetos y que reducen la concentración plasmática impiden la liberación de la hormona antidiurética, produciendo la consiguiente pérdida de líquidos. Esta situación hace que el paciente permanezca en un estado de sed crónica.
-
Polidipsia primaria psicogénica.
Se produce una negación de la sed por parte del paciente y a pesar de ello los volúmenes de ingesta varían entre los 5 y 20 litros. Entre las causas podría ser un daño en el hipotálamo
-
Polidipsia primaria gestacional:
Se produce una ausencia renal a la ADH y una reducción en los niveles de oxitocina OX. Este efecto podría deberse a la liberación de vasopresinasa durante el embarazo.
TEMA 9: PROCESOS SUEÑO-VIGILIA
Introducción.
Los mamíferos, así como la mayoría de las aves alternan a lo largo de sus vidas dos comportamientos: uno de actividad y otro de quietud, con consciencia reducida y recuerdo limitado.
El sueño es un proceso motivacional, pues tarde o temprano los organismos nos vemos forzados a dormir
Estadios o fases del sueño: sueño de ondas lentas y sueño REM.
El cuadro de actividad eléctrica de una persona despierta, activa, es una mezcla desincronizada de ondas, con frecuencias de altas y de baja amplitud. Cuando nos relajamos aparecen unos ritmos diferentes con ondas regulares de frecuencias bajas y alta amplitud (ritmos alfa).
-
Fase 1: a medida que empezamos a tener sueño la amplitud del ritmo alfa decrece y los EEG muestran ondas de frecuencia irregular. Dura unos minutos
-
Fase 2: es definida por unas ondas de alta frecuencia y poca amplitud. dura entre 5 y 15 minutos. Aparecen unas ondas llamadas complejos K que aparecen con la finalidad de que el sujeto no se despierte con el ruido.
-
Fase 3: las ondas comienzan a ganar en amplitud y descienden en frecuencia, son los ritmos delta. Los movimientos de los ojos y del cuerpo están ausentes.
-
Fase 4: es la más profunda del sueño, con ondas EEG de gran amplitud y baja frecuencia.
A partir de aquí el sueño vuelve a ascender a las etapas anteriores, llegando a la fase dos y entrando de repente en un período de sueño REM, con ritmos beta, rápidos e intensos (frecuencias altas) con frecuentes movimientos de los ojos. La respiración y la tasa cardiaca se hace más rápida e irregular. Desciende la temperatura corporal. Se experimentan los sueños.
Características del sueño REM:
-
Registros corticales rápidos, desincronizados
-
Cuando se produce una privación e sueño los sujetos deben recuperarlo poco a poco.
-
Está consolidado en especies desarrolladas: aves y mamíferos.
-
Es uno de los mas profundos junto con la fase 4 de ondas lentas
-
Predomina al nacer y se va reduciendo
-
Nos quedamos bloqueados muscularmente
-
Se producen las ensoñaciones
-
Su comienzo va precedido de las ondas PGO (ponto-geniculadas-occipital). Se encuentran asociadas a centros visuales, preceden a los movimientos oculares que se producen en REM
Anatomofisiología del sueño
-
Un punto de vista considera al sueño como algo pasivo, surge cuando el nivel de vigilancia disminuye. La clave sería estar despierto, y cuando no sucede aparece el sueño. Factores como la acumulación de toxinas resultantes de la actividad facilitan dicho decaimiento.
-
El otro punto de vista considera las teorías como algo activo, pues hay sistemas que expresamente se encargan de que durmamos, por lo tanto el sueño es algo activo y no un mero resultado.
Las estructuras del tronco cerebral son las mas relevantes en el sueño. Se examinó en gatos la actividad eléctrica cortical después de producir lesiones por debajo del tallo cerebral (encéfalo aislado). Sus electroencefalogramas mostraron signos de vigilia y sueño.
Cuando las lesiones se realizaban a nivel del mesencéfalo (preparaciones de cerebro aislado) los animales aparecían en situación de sueño profundo (ondas delta).
Magoun y Moruzzi descubrieron que si estimulaban eléctricamente la parte anterior del tronco los animales se despertaban, los ritmos lentos se hacían rápidos, la zona estimulada fue la formación reticular, su lesión produce la muerte.
Posteriormente dichos autores seccionaron el tronco a nivel medio-pontino (a nivel de la protuberancia), los animales no dormían el 90% del tiempo, lo que podría interpretarse como la interrupción del centro activador del sueño, además, parece demostrar que por encima de la lesión (a nivel rostral de la protuberancia) se encuentra el centro responsable de la actividad(locus cerúleos), por debajo, las estructuras implicadas en el sueño(núcleo del tracto solitario)
¿y que pasa con el sueño REM? parece haberse encontrado una sección de la protuberancia que consta de unos núcleos que rodean al pedúnculo cerebeloso: el núcleo latero dorsal y núcleo pedúnculo pontino, ambos son colinérgicos. Cuando se destruyen se bloquea REM. Además este sistema también organiza las siguientes manifestaciones:
-
Desarrollo de ondas PGO, actuando sobre las zonas pontinas, geniculado lateral y occipital.
-
Atonía muscular a través de las vías descendentes, que van del bulbo a las motoneuronas de la médula, impidiendo la aparición de respuesta motora, si existe lesión en esta vía se realizan movimientos acordes con lo que se sueña.
-
También organizan los movimientos oculares rápidos actuando sobre los colículos superiores.
Estudios más recientes resaltan la importancia de estructuras diencefálicas en el control y regulación de estos procesos, la estimulación eléctrica de baja frecuencia en el núcleo intralaminar del tálamo en conejos indujo al sueño de ondas lentas.
El papel del tálamo también es de vital importancia, existe una enfermedad mortal llamada insomnia familiar fatal, en la que tras sufrir una etapa de insomnio, mueren. Tras la autopsia se observa degeneración y pérdida neuronal talámica. Su explicación parece residir en la presencia de una proteína anormal en el cerebro.
También se han observado alteraciones en el sueño en personas con disfunciones hipotalámicas. Por ejemplo la encefalitis letárgica. Producida por lesiones extensas del hipocampo que producían un estado de sueño prolongado . Tal explicación es que durante el sueño no REM se produce una activación del área preóptica ventrolateral, esta región contiene neuronas gabaérgicas inhibitorias que proyectan al hipotálamo posterior (a los núcleos tuberomamilares) que poseen células histaminérgicas que proyectan al tálamo y córtex.
Neuroquímica del sueño
-
Pieron comprobó que al realizar trasfusiones de fluido cereboespinal de animales privados de sueño a sujetos normales no privados, estos dormían. Lo que hizo que muchos investigadores pensasen en la existencia de factores endógenos.
-
En 1975 Pappenheimer identificó un péptido, el factor S como posible promotor de sueño de ondas lentas al ser administrado intraventricularmente. Esta sustancia no es producida por los mamíferos, sino por bacterias degradadas por el sistema inmunológico, esto hizo pensar que sería posible que el factor S hace que el sueño aparezca como una respuesta del sistema inmunitario para combatir una posible infección.
-
El factor más estudiado como promotor de sueño ha sido el DSIP (péptido inductor del sueño Delta). Se ha obtenido de la circulación cerebral y tiene efectos sobre el sueño cuando se administra intravenosa o intramuscularmente. Para saber cómo llega al cerebro desde el sistema inmunitario se ha estudiado en siameses, en todos los casos el sueño aparecía independientemente de si la otra persona estaba despierta o dormida.
-
Otra sustancia ha sido implicada en el sueño, la adenosina, su efecto inductor del sueño puede deberse a la inhibición de varios neurotransmisores -como la noradrenalina- implicados en el arousal. El efecto de la cafeína podría ser debido a la inhibición de los receptores de la adenosina.
El hecho de que los niveles de algunos neurotransmisores varíen a lo largo del día ha sugerido que el comienzo del sueño podría estar controlado por cambios en la actividad de diferentes sistemas de neurotransmisor.
-
Serotonina y noradrenalina: actúan conjuntamente, se han observado niveles elevados de ambos en el locus cerúleos durante la vigilia, y se van reduciendo durante el sueño de ondas lentas hasta desaparecer en el sueño REM
-
Acetilcolina: sus niveles aumentan antes y durante REM. Fármacos como la fisostigmina o carbacol provocan la aparición de REM (son agonistas), y antagonistas como la atropina o escopolamina reducen este tipo de sueño
-
Dopamina e histamina: durante la actividad la histamina es elevada, así como las enzimas responsables de su síntesis y degradación. La facilitación de la dopamina incrementa la actividad locomotora (anfetaminas y metilfenidato)
-
Benzodiacepinas: actúan sobre los complejos K facilitando el sueño (triazolan)
TEMA 10: BASES BIOLÓGICAS DE LA HABITUACIÓN Y CONDICIONAMIENTO CLÁSICO
Habituación y sensibilización. Mecanismos neurobiológicos
El aprendizaje es el proceso por el cual se adquieren conocimientos sobre el mundo. La memoria es el almacenamiento de dicho conocimiento.
Hay que diferenciar entre aprendizaje explícito o consciente entre aprendizaje mecánico, automático o implícito.
Eric Kandel estudió la habituación en un caracol marino, la Aplysia, con un pequeño de neuronas de gran tamaño.
La Aplysia tiene una branquia sobre la que se sitúa el manto y sifón. Cuando estimulamos táctilmente el manto o el sifón se produce un reflejo defensivo, la retirada de la branquia, mediante estímulos repetidos este reflejo se habitúa.
Con mayor detalle, diremos que como respuesta ante un estímulo novedoso, las neuronas sensoriales que lo inervan generan potenciales postsinápticos excitatorios en las interneurona y motoneuronas. Estos potenciales conducen a un potente reflejo de retirada de la branquia. Si el estímulo se presenta repetidamente, los potenciales sinápticos producidos por las neuronas sensoriales en las interneuronas y motoneuronas se vuelven progresivamente menores. La disminución parece obeceder a una menor cantidad de Glutamato en la terminal presináptica, esta inactivación puede venir por una inhibición de los canales de calcio en la terminal presináptica, debido a la estimulación repetida. En definitiva, la habituación produce depresión monosináptica
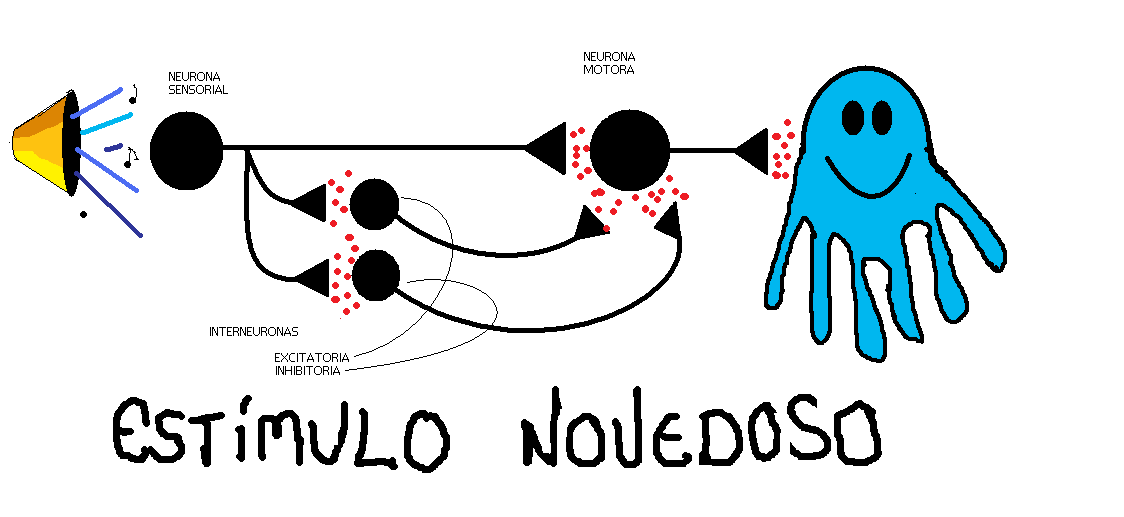
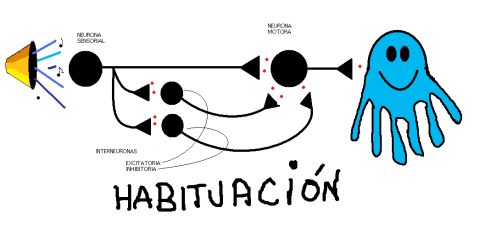
Existe otro aprendizaje, la sensibilización, en este caso, el animal, después de aprender que un estímulo es nocivo, responde más rápidamente frente a otros estímulos, implica facilitación heterosináptica, es decir, el estímulo sensibilizante activa un grupo de inerneuronas que forman sinápsis con las neuronas sensoriales (axoaxónicas). La serotonina y otros neurotransmisores se liberan por neuronas facilitadoras que provocan que permanezcan abiertos por más tiempo los canales de calcio y por lo tanto aumentan los niveles del neurotransmisor.
Cuando la habituación y la sensibilización se establecen firmemente se convierten en aprendizajes a corto plazo, se ha observado que no solo cambia el mecanismo químico sino también se producen cambios estructurales. Estos cambios pueden atribuirse a genes y proteínas pues por ejemplo los animales sensibilizados tienen dos veces mas terminales presinápticas que los animales no entrenados. Tales cambios son característicos de la sensibilización a largo plazo, por otra parte la habituación a largo plazo conduce a la pérdida de conexiones sinápticas entre las neuronas sensoriales y las neuronas motoras
Condicionamiento clásico en invertebrados, mecanismos celulares.
Kandel presentaba un estímulo táctil que evocaba pocas respuestas, a continuación proporcionaba un shock eléctrico, tras un número breve de repeticiones el tacto por sí mismo provocaba un mayor número de respuestas.
Dentro de esta asociación entre EC y EI es muy importante la contigüidad temporal, pues para que se de condicionamiento el EC debe preceder al EI dentro de al menos un intervalo de 0,5 seg. ¿Por qué?
El EI en la cola produce la facilitación de las inetrneuronas que tienen conexiones axoaxónicas con las neuronas sensoriales del EC (sensibilización) este hecho conduce a la facilitación presináptica. Sin embargo, si los dos estímulos se están secuenciados, las neuronas sensoriales comienzan a disparar como respuesta al EC, pero en ese momento las interneuronas activadas por el EI producen una mayor facilitación presináptica en las neuronas sensoriales (mayor liberación de nt). Por el contrario la facilitación no se potencia si las interneuronas se activan antes de que comiencen a activarse las neuronas sensoriales.
Condicionamiento clásico en mamíferos
Uno de los aprendizajes mas estudiados ha sido el condicionamiento nictitante: consiste en emparejar un estímulo condicionado (sonido) con un estímulo incondicionado (soplo de aire en el ojo) que produce un despliegue ede la membrana ocular que tienen algunos mamíferos, tras varios emparejamientos se provoca la respuesta ante el EC solo.
Thompsom comprobó que estructuras eran las responsables, encontró una correlación entre la actividad hipocampal y el aprendizaje, aunque si se lesionaba el hipocampo el aprendizaje nictitante seguía produciéndose. Por lo tanto fijó sus observación en uno de los núcleos profundos del cerebelo, el interpósito, la actividad celular de este núcleo se modificaba con la conducta, y las lesiones en esta región interrumpían el aprendizaje.
El hipocampo está mas relacionado con procesos complejos del condicionamiento, formaría parte de aprendizajes como el condicionamiento de huella donde los dos estímulos están presente pero el EC aparece después del EI, sería relevante, pues para asociar dos estímulos cuando uno no está presente.
TEMA 11. ETOLOGÍA: APRENDIZAJE ESPECÍFICO DE LAS ESPECIES. APRENDIZAJE INTEROCEPTIVO.
Aprendizaje interoceptivo. Características.
El hecho de que cualquier aprendizaje se puede explicar por el condicionamiento clásico fue rechazado por una serie de estudios que existes otros aprendizajes, entre ellos se encuentra el aprendizaje interoceptivo: aversivo-gustativo o toxifobia. Es un aprendizaje que se encuentra determinado genéticamente y se encuentra presente en todas las especias, está supeditado a una base biológica. Consiste básicamente en evitar la ingesta de sustancias que han sido emparejadas con una situación nociva de origen visceral (tóxica). Un caso similar es el de la quimioterapia.
El paradigma experimental más utilizado en este tipo de aprendizaje consiste en implantar en los animales unas fístulas intragástricas que ponen el sistema gastrointestinal al alcance de cualquier manipulación exterior. El procedimiento que le sigue es el siguiente:”una vez recuperado el animal de la intervención quirúrgica se le ofrece un estímulo gustativo con un sabor determinado, que irá seguido tras una demora de 15 minutos de la administración de un producto aversivo (Cloruro de Litio, Cloruro Sódico…). Al día siguiente se le presenta otro sabor diferente y posteriormente, después de 15 minutos, se le administra suero fisiológico 0,9% (sustancia neutra). En el tercer día se le presentan al animal dos estímulos gustativos para observar su preferencia”.
Los animales experimentales suelen elegir el sabor que había sido emparejado con el suero y aborrecen el otro y si solo tienen que tomar esa sustancia, prefieren morir
Existe otro procedimiento experimental en el cual el animal tiene implantadas dos fístulas intragástricas: “En el primer día de experimento se le presentan dos estímulos a la vez, de forma que pueda elegir entre ellos. En el momento que prueba uno de ellos se le administra intragástricamente y de forma simultánea una sustancia neutra (suero fisiológico al 0,9%) mientras que si toma del otro sabor se le administra simultáneamente una sustancia aversiva (sulfato de cobre). Tras varias repeciciones el animal aprende y se decanta por el estímulo gustativo que no está asociado con el producto aversivo”. Una vez analizados los resultados se pueden constatar las siguientes características:
-
Se aprende a asociar un sabor con un malestar aunque este se produzca horas después de tomar el alimento, se aprende sin contingencia y sin contigüidad
-
Este tipo de aprendizaje se produce con un solo ensayo
-
En el aprendizaje aversivo-gustativo se asocia un estímulo específico (sabor) con uno gástrico, en el condicionamiento clásico el estímulo puede ser condicionado por igual (equipotencialidad)
-
Puede darse en animales curalizados (paralizados). Por lo tanto, para dicho aprendizaje no es necesario una respuesta motora, como sucede en condicionamiento clásico.
Mecanismos de transmisión al sistema nervioso central.
En algún lugar del SNC se debe producir la convergencia de los estímulos gustativos y viscerales:
-
La información gustativa es transmitida por los diferentes pares craneales (VII, IX, X) hasta el núcleo del tracto solitario, su lugar de enlace en el tronco.
-
La información visceral se transmitirá al SNC de manera más compleja:
-
Habría un mecanismo sensorial rápido capaz de detectar la presencia de estímulos nocivos en el sistema gastrointestinal (nervio vago: con sus tres tipos de receptores -mecanorreceptores, quimioreceptores y osmorreceptores). Si los animales son vagotomizados no son capaces de aprender a corto plazo.
-
También contamos con un sistema más lento, formado por el sistema circulatorio, una vía humoral, sanguínea, que detectan los cambios que se producen en el interior del organismo.
Centros de convergencia cerebral: núcleo del tracto solitario, área postrema y complejo parabraquial.
-
En el caso del aprendizaje a corto plazo, proveniente del Vago, la información llegará al núcleo del tracto solitario, posteriormente llegará al núcleo parabraquial medial
-
En el caso del aprendizaje a largo plazo, la información que proviene del sistema circulatorio tiene su relevo en el área postrema termina en el núcleo parabraquial lateral
No obstante se ha podido observar como ratas descerebradas son incapaces de formar asociaciones necesarias para aprender la aversión al gusto, por lo que la integración en el tallo no parece suficiente para explicar la adquisición y expresión de las aversiones gustativas.
Ciertas estructuras del cerebro anterior (amígdala y córtex insular) parecen implicadas en este tipo de aprendizaje. Las lesiones de la amígdala han mostrado resultados contradictorios, por lo que se podría pensar que si se produce algún efecto puede ser debido al daño de las fibras pertenecientes al córtex insular que transcurren por la amígdala. Se piensa así porque el córtex insular mantiene conexiones recíprocas con el núcleo del tracto solitario y núcleo parabraquial, estudios demuestran la atenuación del aprendizaje aversivo gustativo después de la destrucción de esta región
TEMA 12: PSICOBIOLOGÍA DEL APRENDIZAJE Y LA MEMORIA
Introducción: lóbulo temporal medial.
Penfield en los años 40 logró mediante estimulación eléctrica cortical produjo respuestas en los pacientes que describían en forma de recuerdos coherentes de experiencia anteriores, lo logró estimulando los lóbulos temporales.
HM fue una persona sometida a una extirpación del lóbulo temporal medial, tras lo cual experimentó una gran pérdida de memoria y fue incapaz de tener nuevos recuerdos, por lo tanto tampoco podía tener recuerdos a largo plazo, los recuerdos anteriores a la operación quedaron intactos.
Identificación de los componentes del sistema de memoria: lóbulo temporal medial
Las investigaciones se han centrado en tres campos principales:
-
Estudios con pacientes neurológicos que tienen problemas de amnesia
-
Trabajos experimentales con modelos de amnesia en primates
-
Estudios de lesiones selectivas en ratas.
En 1978 se observó que lesiones extensas (amígdala, hipocampo, córtex perirrinal, parahipocampal y entorrinal, lesión H+A+) del lóbulo temporal medial en monos provocaban graves deterioros en la memoria.
Posteriormente se llevaron a cabo lesiones selectivas en distintas partes del lóbulo temporal medial:
-
Se lesionó el hipocampo y la zona parahipocampal, -lesión H+-, estas lesiones provocaron serios problemas en tareas de memoria.
-
Se lesionó solo el hipocampo,,-lesión H- y mostraron graves alteraciones en la memoria pero menores a las observadas tras las lesiones H+.
-
Los monos sometidos a las lesiones H+A+, obtienen peores resultados que los observados con la lesión H+.
Se realizaron lesiones electrolíticas en el complejo amigdalino, dejando intactas las cortezas vecinas (lesión A), los monos ejecutaban normalmente las tareas de memoria.
El gran deterioro observado en H+A+ puede ser atribuido al daño en las regiones que rodean a la amígdala, pues animales a los que se les lesionó el hipocampo y las cortezas adyacentes (Parahipocampo, Entorrinal y Perirrinal), -lesión H++-, mostraron prácticamente los mismos problemas de memoria que los descritos en las lesiones H+A+.
Las lesiones hipocampales o de estructuras relacionadas perjudicaron en una gran variedad de tareas de memoria: espacial, asociación…conclusiones.
-
El hipocampo y las estructuras relacionadas son importantes en la memoria declarativa.
-
Los déficits asociados a lesiones hipocampales, pueden ser incrementados por daño adicional en regiones y tractos relacionados anatómicamente.
Formación hipocampal y memoria.
En ausencia de hipocampo todos los aprendizajes y eventos que aparecen después de la lesión se pierden completamente. Trabajos recientes en monos con lesiones bilaterales en la formación hipocampal (Lesión H+) parecen confirmar estas ideas. Estos animales fueron entrenados preoperatoriamente en una tarea de discriminación de pares de objetos. El entrenamiento de cada uno de los 20 pares comenzó 16, 12, 8, 4 y 2 semanas antes de la operación. Dos semanas después de la operación se evaluó la memoria del animal para la presentación de cada uno de los 100 pares de objetos. El resultado más importante fue que los monos operados recordaron mejor aquellos objetos aprendidos mucho antes de la operación que los aprendidos recientemente, estos resultados fueron opuestos al grupo control. Estos datos indican que el papel del Hipocampo en la memoria es únicamente temporal. La memoria sería gradualmente reorganizada y consolidada con el paso del tiempo en otro lugar, en el neocórtex.
Modulación neuroquímica de la memoria. Potenciación a largo plazo (PLP)
Lomo y Bliss (1966) descubrieron que la estimulación intensa de axones de regiones hipocampales producen un incremento duradero (a largo plazo) en la magnitud de los potenciales postsináptico excitadores con las células postsinápticas, este incremento se denomina potenciación a largo plazo (PLP).
Si seguimos la norma de Hebb, donde si A activa a B y éste se encuentra ya activado (despolarizado) entonces A se vuelve fortalecido (fortalecimiento sináptico). Kelso, Ganong, y Brown encontraron que si despolarizaban artificialmente neuronas CA1 y entonces estimulaban los axones que formaban sinapsis con ellas, éstas se volvían más fuertes, si la estimulación y la despolarización ocurrían en tiempos diferentes, el efecto no se producía. Esto quiere decir que para que se dé la PLP se require: activación de la sinapsis A y despolarización de la neurona postsináptica B.
A nivel fisiológico, la explicación reside en las características del receptor NMDA, se encuentran en varias regiones del hipocampo.
El NMDA controla un canal de calcio (interacción alostérica). En condiciones normales, cuando el glutamato (neurotransmisor que actúa por el NMDA), estimula a éste no se produce al entrada de iones de calcio en la terminal, ya que el canal NMDA se encuentra bloqueado por el magnesio. Pero si la membrana postináptica es despolarizada y en ese momento actúa el glutamato, el magnesio es expulsado del canal y éste se abre al calcio. Por lo tanto, el calcio entra en la célula a través de los canales controlados por NMDA únicamente cuando el glutamato está presente y la membrana postináptica ha sido despolarizada.
La fuerte evidencia proviene de la investigación con fármacos que bloquean estos receptores. La administración de AP5 impide la PLP en CA1 y en el Giro Dentado, dos regiones importantes del hipocampo. Sin embargo este efecto no se observa cuando la PLP ya ha sido establecida.
¿Qué mecanismo será el responsable para el incremento en el fortalecimiento sináptico que ocurre durante la potenciación a largo plazo? Alternativas:
-
La PLP se produce como consecuencia de un mecanismo presináptico que actúa liberando más neurotransmisor, otros piensan que es un sistema postsináptico, mediante la existencia de un mayor número de receptores. De todas formas sería factible que la PLP implique cambios tanto pre como post.
-
Algunos autores indican que la PLP es crucialmente postsináptica. A raíz de la entrada de iones de Calcio en la terminal, se activarían una serie de enzimas: proteína Kinasa C, Tipo II Calcio Calmodulina Kinasa y Tirosina Kinasa. Estas enzimas quizás incrementan la sensibilidad de los receptores postsinápticos, eleven el número de receptores para el glutamato o lleven a cabo cambios presinápticos.
¿Como un proceso que ocurre en las espinas dendríticas de la neurona postsináptica puede causar modificaciones a nivel presináptico?
Una explicación sería el descubrimiento de unas moléculas simples (gases) como el óxido nítrico y el monóxido de carbono que podrían llevar mensajes de una célula a otra (post a pre) promoviendo la mayor liberación de neurotransmisor.
Para saber si el proceso de potenciación a largo plazo puede explicar la consolidación y el almacenamiento de la memoria sería interesante comprobar si los métodos que bloquean la PLP también impiden el aprendizaje y memoria de una serie de tareas. Para ello, Richard Morris inyectó un bloqueador de los receptores NMDA en el hipocampo de ratas que habían sido previamente entrenadas en una piscina, posteriormente no recordaron la localización de la plataforma sumergida.
Otros estudios que han confirmado la importancia de la PLP en la memoria han sido los desarrollados por Alsacino Silva los cuales producen una serie de mutaciones en la enzima Calmodulina Kinasa II provocando una alteración en el aprendizaje espacial de los animales.
Descargar
| Enviado por: | Jaen |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.