Salud
Neuroanatomía
ANATOMIA HUMANA
TRABAJO SOBRE NEUROANATOMIA
EMBRIOLOGIA DEL SISTEMA NERVIOSO
Introducción
Los procesos de inducción, migración y diferenciación celular que se llevan a cabo durante la formación del tejido nervioso generan un sistema altamente organizado capaz de proporcionar al nuevo ser una eficiente red de comunicación con gran respuesta adaptativa y con la peculiaridad de responder autónomamente a estímulos físicos y químicos originados tanto en el medio interno como en el circundante (exitabilidad). De esta manera, el sistema nervioso central (SNC) permite integrar y controlar todas las regiones del organismo.
Si se observa la evolución de las especies, la centralización de la información es uno de los principios básicos de la organización de los seres vivos, y es el SNC el encargado de asumir tales funciones. Un conocimiento básico de la embriología ayuda a comprender de mejor manera las intrincadas interrelaciones de los distintos componentes del SNC.
Desarrollo del Tubo Neural
El sistema nervioso comienza su desarrollo embriológico en la tercera semana (embrión de unos 1,5 mm.) en la porción medial de la región dorsal a partir del cuarto par de somitos (entre la membrana bucofaríngea y el nodo primitivo).
Los procesos de formación de la placa neural, pliegues neurales, y desarrollo del tubo neural se agrupan en el concepto de neurulación. Este período abarca desde el proceso de inducción notocordal hasta el cierre del neuroporo caudal. Durante el proceso de neurulación el embrión suele llamarse néurula. He aquí una breve descripción de estos procesos:
Al comenzar la tercera semana, la notocorda en desarrollo y el mesodermo adyacente estimulan al ectodermo que está encima de ellos. Este complejo proceso de inducción notocordal hace que tejido ectodérmico (neuroectodermo) se engruese, formándose así la placa neural. Es posible que el engrosamiento del neuroectodermo tenga relación con la polimerización de microtúbulos en el citoplasta de las células nerviosas en formación. Actualmente, se han identificado varios tipos de moléculas que actúan como señales en procesos inductivos del SNC, algunas de ellas pertenecen a la familia del factor transformador del crecimiento Beta (FTC Beta), otras al grupo de los factores de crecimiento de los fibroblastos (FCF). Existen otras moléculas como el ácido retinoico o cierto tipo de neurotransmisores que al encontrarse en gradientes de concentración producen respuestas celulares importantes. La inducción neural trae como consecuencia una sobreproducción inicial de células nerviosas. Se ha demostrado que a tal período prosigue otro de muerte celular (muerte celular programada o apoptosis por fragmentación celular) que determina la cantidad total de neuronas que el individuo tendrá durante su vida. Tal número alcanza la impresionante cifra de un billón.
Una vez completado el proceso inductivo, la placa neural se alarga desde su sitio de origen craneal al nodo primitivo hasta la membrana bucofaríngea. Alrededor del 18º día de desarrollo los bordes laterales de la placa neural se elevan y forman los pliegues neurales; la porción media entre los pliegues neurales forma el surco neural. Hacia el final de la tercera semana los pliegues neurales se elevan aún más, se acercan y se fusionan irregularmente en la línea media formando el tubo neural. La fusión empieza en la región cervical y sigue hacia cefálico y caudal. Mientras ocurre la fusión, los bordes libres del ectodermo superficial se separan del tubo neural. Posteriormente, ambos bordes se unen y forman una capa continua en la superficie que dará origen al epitelio epidérmico.
Debido a que la fusión de los pliegues neurales no ocurre simultáneamente a lo largo de ellos, la luz del tubo neural comunica con la cavidad amniótica en sus extremos cefálico y caudal a través de los neuroporos craneal (anterior) y caudal (posterior). El cierre del neuroporo craneal se realiza en ambas direcciones desde el sitio inicial de cierre en la región cervical o desde otro que se origina un tiempo después en el prosencéfalo que avanza en dirección caudal. Este cierre ocurre al 25º día (período 18-20 somitos). El neuroporo caudal se cierra el 27º día (período de 25 somitos). El cierre de ambos neuroporos coincide con el establecimiento de la circulación sanguínea hacia el tubo neural.
Mientras los pliegues neurales se acercan a la línea media para fusionarse, un grupo de células neuroectodérmicas ubicadas en la cresta de cada pliegue (cresta neural) pierden su afinidad epitelial con las células de la vecindad. La migración activa de las células de la cresta neuraldesde las crestas hacia el mesodermo adyacente transforma el neuroectodermo en tejido mesenqumático que forma una masa aplanada e irregular que rodea al tubo neural. Este grupo celular dará origen a un conjunto heterogéneo de tejidos de gran importancia: ganglios de la raíz posterior, ganglios autónomos, ganglios de los pares craneales V, VII, IX, X, células de Schwann, las leptomeninges (aracnoides y piamadre), melanocitos, médula suprarrenal, odontoblastos. En consecuencia, el tubo neural será el que se convertirá por diferenciación en encéfalo y médula espinal, mientras que las crestas neurales formarán la mayor parte del sistema nervioso periférico (SNP) y parte del autónomo (SNA).
Luego del cierre completo del tubo neural, comienza el desarrollo de la región caudal del tubo (segmentos sacros bajos y coccígeo) mediante procesos de canalización y diferenciación regresiva. Entre el 28º y 32º día, un conjunto de células indiferenciadas al final del tubo neural desarrolla una serie de pequeñas vacuolas que posteriormente toman contacto con el canal central. Es frecuente la presencia de innumerables canalículos que pudiesen ser causas de posteriores defectos. Este proceso de canalización prosigue hasta la séptima semana. En el periodo siguiente ocurre la regresión de muchas células de la masa caudal. Como remanentes de este proceso de diferenciación regresiva quedan el ventrículo terminal y el filum terminale. El extremo cefálico del tubo neural se dilata y origina 3 vesículas encefálicas primarias:
-Prosencéfalo (cerebro anterior)
-Mesencéfalo (cerebro medio )
-Rombencéfalo (cerebro posterior)
El tercio caudal del tubo se alarga y su diámetro se acorta para formar la médula espinal.
El neurocele se estrecha y pasa a formar el canal central (del epéndimo) de la médula espinal que se continúa con la cavidad de las vesículas encefálicas. La cavidad del rombencéfalo es el Cuarto ventrículo, la del diencéfalo el Tercer ventrículo y la de los hemisferios cerebrales los Ventrículos laterales. Tercer y cuarto ventrículos se comunican por la luz del mesencéfalo que se torna estrecha y origina el Acueducto cerebral (de Silvio). Los ventrículos laterales se comunican con el Tercer ventrículo por los agujeros interventriculares (de Monro).
Médula Espinal
Luego de ocurridos los sucesos de neurulación, el tubo neural forma una estructura totalmente separada de la cavidad amniótica cuya pared está constituida por células cilíndricas que la recorren formando un epitelio pseudoestratificado y que están conectadas por complejos de unión (zónula occludens, zónula adherens y desmosomas) hacia el neurocele. Durante el período de surco neural incluso hasta después de cerrarse el tubo neural, estas células proliferan para formar la Capa neuroepitelial (neuroepitelio). Esta capa da origen a todas las neuronas y neuroglias (astrocitos y oligodendrocitos) de la médula espinal. Durante la quinta semana, las células neuroepiteliales proliferan y producen un aumento en longitud y diámetro del tubo neural. Además, es posible observar cambios en la conformación de los diferentes elementos intracelulares, como por ejemplo, modificaciones en la morfología del núcleo o la presencia de un mayor número de ribosomas asociados al retículo endoplásmico consecuencia del considerable aumento en la actividad neurosecretora. Estas células denominadas Neuroblastos (células nerviosas primitivas) migran a la periferia y se organizan en una nueva estructura: la Capa del manto, la que posteriormente constituirá la sustancia gris de la médula espinal.
Las prolongaciones axonales de las neuronas de la capa del manto migran a la periferia y forman los fascículos nerviosos de la Capa marginal. Al mielinizarse estas fibras nerviosas, la capa toma un aspecto blanquecino y constituye la sustancia blanca de la médula espinal.
Gran parte de los glioblastos (células de sostén primitivas) deriva del neuroepitelio una vez que este ya ha dado origen a los neuroblastos. Los glioblastos emigran desde la capa neuroepitelial hacia las capas marginal y del manto para allí diferenciarse en Astrocitos tipo I (aquellos que envían prolongaciones a la piamadre y a los vasos sanguíneos encefálicos para formar la barrera hematoencefálica) y tipo II (que toman contacto con los nodos de Ranvier de los nervios mielínicos del SNC y suelen encapsular las sinapsis químicas). Los astrocitos tienen importantes funciones en el SNC: 1) Forman la barrera hematoencefálica que protege al SNC de cambios bruscos en la concentración de iones del líquido extracelular y de otras moléculas que pudiesen interferir en la función neural. 2) Eliminan K+, glutamato y ácido ,-aminobutírico del espacio extracelular. 3) Son importantes almacenes de glucógeno; realizan glucogenólisis al ser inducidos por norepinefrina o péptido intestinal vasoactivo (VIP).
La célula de oligodendroglia es posible que derive de los glioblastos. En la capa marginal forma las vainas de mielina que rodean a los axones. Los oligodendrocitos satélites son aquellos que rodean los cuerpos celulares y regulan el ambiente bioquímico que rodea la neurona.
Alrededor del cuarto mes aparecen las células de microglia. Derivan del mesénquima circundante y se caracterizan por ser pequeñas y muy fagocíticas. Llegan a la sustancia blanca y gris del SNC luego de la aparición de los vasos sanguíneos.
Cuando las células neuroepiteliales dejan de producir neuroblastos y glioblastos, se diferencian las células ependimarias que revisten el canal central de la médula espinal.
Desarrollo de las placas basales, alares, del techo y del piso: La multiplicación los neuroblastos de la capa del manto a cada lado del tubo neural origina unos engrosamientos en la región ventral y dorsal: 1)Las placas basales (engrosamiento ventral) incluyen los somas de las motoneuronas que posteriormente constituirán las astas anteriores de la médula espinal. 2)Las placas alares (engrosamientos dorsales) corresponden a regiones sensitivas que se diferenciarán en las astas posteriores de la médula espinal. El crecimiento de las placas alares origina el tabique medio posterior. Al sobresalir ventralmente las placas basales se forma el tabique medio anterior, mientras tanto se desarrolla la fisura mediana anterior en la superficie anterior de la médula espinal. El surco limitante delimita ambas placas, y de esta manera también separa las regiones motoras de las sensitivas. Las regiones dorsal (placa del techo) y ventral (placa del piso) en la línea media del tubo neural no poseen neuroblastos y constituyen vías para fibras nerviosas que cruzan la médula espinal de un lado al otro.
Entre las astas ventral y dorsal de los segmentos torácicos hasta el segundo o tercero lumbar de la médula espinal se acumulan neuronas que formarán el asta lateral o intermedia, que contiene neuronas del Sistema nervioso autónomo.
Las meninges espinales y las modificaciones durante la formación de la médula espinal: El tejido mesenquimático (esclerotoma) que rodea el tubo neural se condensa para formar la meninge primitiva. Su capa externa se engrosa para originar la duramadre. La capa interna permanece delgada para formar las leptomeninges (aracnoides y piamadre). Al unirse los espacios llenos de líquidos que existen entre las leptomeninges, se forma el espacio subaracnoídeo. El origen de la aracnoides y piamadre a partir de una capa única explica la existencia de las trabéculas aracnoideas que pasan entre ellas.
Como resultado del desarrollo del aparato locomotor durante el cuarto mes, además de la adición de neuronas motoras y sensitivas, la médula espinal se ensancha en las regiones cervical y lumbar formando los engrosamientos cervical y lumbar.
Al tercer mes, la médula espinal se extiende a lo largo del canal vertebral del embrión y los nervios espinales atraviesan los agujeros intervertebrales a nivel de su origen. Poco después, la columna vertebral y la duramadre se alargan más rápido que el tubo neural ocasionando que el extremo terminal de la médula se desplace a niveles más altos. A los seis meses de vida intrauterina alcanza la primera vértebra sacra, y ya en el neonato su extremo está a nivel de L3. Debido a este crecimiento desproporcionado, los nervios raquídeos tienen una dirección oblicua desde su segmento de origen en la médula espinal hasta el nivel correspondiente de la columna a nivel coccígeo.
En el adulto, la médula espinal termina a nivel L2 (esta es una medida promedio, ya que el extremo medular puede estar tan alto como T12 o tan bajo como L3). Debajo, una prolongación filiforme de la piamadre forma el filum terminale que se adosa al periostio de la primera vértebra coccígea y señala la línea de regresión de la médula espinal embrionaria. Las fibras nerviosas bajo el extremo inferior de la médula espinal forman la Cauda equina, cuya denominación se debe a su semejanza a la cola de caballo. Cuando se extrae LCR por una punción lumbar, la aguja se introduce en un nivel lumbar bajo respetando así el extremo terminal de la médula espinal.
Encéfalo
Las estructuras encefálicas aparecen luego de ocurridos cuatro procesos básicos: (a) proliferación neuronal (b) migración (c) período de organización, el cual se desarrolla hasta el nacimiento una vez establecido el patrón de funcionamiento de las diferentes regiones encefálicas, y (d) mielinización.
Durante la cuarta semana, después del cierre de los neuroporos, el extremo cefálico del tubo neural craneal al cuarto par de somitos se dilata considerablemente y aparecen las tres vesículas encefálicas primarias a partir de las cuales se origina el encéfalo: Prosencéfalo (cerebro anterior), Mesencéfalo (cerebro medio) y Rombencéfalo (cerebro posterior). Simultáneamente se están formando dos plegamientos: el pliegue cervical, en la unión del rombencéfalo y médula espinal y el pliegue cefálico en el mesencéfalo. El mesencéfalo dista del rombencéfalo por un surco: el istmo del rombencéfalo (de His).
Durante la quinta semana el prosencéfalo y rombencéfalo se dividen en dos vesículas secundarias: El prosencéfalo origina (1) el Telencéfalo (cerebro terminal) que consta de una parte media y dos evaginaciones laterales (hemisferios cerebrales primitivos) (2) el Diencéfalo, que presenta la evaginación de las vesículas ópticas. El rombencéfalo formará finalmente (1) el Metencéfalo, que constituirá la protuberancia y el cerebelo (2) el Mielencéfalo, el futuro bulbo raquídeo. El límite metencéfalo-mielencéfalo queda definido por el pliegue protuberancial. Este pliegue se origina debido al crecimiento desigual del rombencéfalo dando lugar a un adelgazamiento de su techo.
Al principio, el encéfalo tiene su estructura básica muy similar a la médula espinal, sin embargo, debido a la aparición de los pliegues y surcos encefálicos se producen variaciones considerables en la disposición de los diferentes elementos. En general, las placas alares y basales del rombencéfalo y mesencéfalo se encuentran bien definidas, en cambio, en el prosencéfalo las placas alares están acentuadas y las basales en plena regresión.
ROMBENCEFALO
Está formado por: (1) Mielencéfalo (2) Metencéfalo
MIELENCEFALO
Es la vesícula encefálica más caudal y se diferencia en el bulbo raquídeo (médula oblonga). Sus paredes laterales sufren cierta eversión tal como se abren las conchas de una almeja, sin embargo, su estructura general es bastante parecida a la médula espinal.
Los neuroblastos de las placas alares migran a la capa marginal en dirección ventrolateral para formar los núcleos olivares. Ventralmente, las fibras corticospinales que descienden desde la corteza cerebral (giro precentral) forman las denominadas pirámides. El pliegue protuberancial hace que las paredes bulbares laterales se desplacen lateralmente y que la placa del techo se extienda y adelgace considerablemente. Como consecuencia, la cavidad del mielencéfalo (futuro IV ventrículo) toma forma romboide y los núcleos motores pasan a ubicarse medialmente a los núcleos sensitivos.
Las placas alares y basales están bien definidas. La placa basal contiene 3 grupos de núcleos motores: (1) Eferente somático o medial (2) Eferente visceral especial o intermedio (3) Eferente visceral general o lateral. Estos tres grupos originan los núcleos motores de los nervios craneales IX, X, XI y XII que se ubican en el piso del cuarto ventrículo medial al surco limitante. La placa alar contiene tres grupos nucleares sensitivos: (1) Aferente somático o lateral (2) Aferente visceral especial o intermedio (3) Aferente visceral general o medial. Estos grupos neuronales forman los núcleos sensitivos de los nervios craneales V, VII, VIII, IX y X y los núcleos gracilis y cuneatus.
METENCEFALO
Incluye la región ubicada entre el pliegue protuberancial y el istmo del rombencéfalo. La porción metencefálica ventral más una contribución celular de la región alar del mielencéfalo originan el Puente (protuberancia), mientras la región posterior conforma el cerebelo. El puente forma una importante vía nerviosa entre la médula espinal y las cortezas cerebral y cerebelosas. Por otra parte, el cerebelo es un centro de coordinación de postura y movimientos. La cavidad del metencéfalo forma la parte superior del futuro IV ventrículo.
La formación del pliegue protuberancial produce el distanciamiento de las paredes laterales del puente y la extensión de la sustancia gris del piso del IV ventrículo.
Los neuroblastos de las placas basales constituyen tres columnas de núcleos motores: (1) Eferente somático medial (2) Eferente visceral especial (3) Eferente visceral general. Ellos originan los núcleos motores de los pares V, VI y VII. La capa marginal de las placas basales se expande y sirve de puente a fibras que conectan la médula espinal con las cortezas cerebral y cerebelosas; esto explica el nombre de "puente".
Las placas alares poseen 2 grupos sensitivos: (1) Aferente somático lateral (2) Aferente visceral general. Ellos constituyen el núcleo sensitivo principal del n.trigémino, el núcleo sensitivo del VII par y los núcleos vestibular y coclear del VIII par. Los núcleos pontinos se originan en las placas alares del metencéfalo y mielencéfalo.
Cerebelo: Cada placa alar se curva en su región dorsolateral en dirección medial para formar los labios rómbicos. Estos labios aumentan de tamaño, se proyectan caudalmente sobre la placa del techo del IV ventrículo y se fusionan en la línea media. En la zona inferior del metencéfalo están muy separados. La compresión cefalocaudal de los labios producto de la exageración del pliegue protuberancial forma la placa cerebelosa que se superpone al puente y al bulbo raquídeo. En el embrión de 12 semanas se observa una parte media (vermis) y dos laterales (hemisferios). Inicialmente, la placa cerebelosa consta de las capas neuroepitelial, del manto y marginal, pero luego algunas células neuroepiteliales emigran a la superficie cerebelosa a formar la capa granulosa externa que consta de una zona proliferativa superficial. Al sexto mes, la capa granulosa externa ya ha producido células granulosas, células en cesto y células estrelladas que contactan con células de Purkinje aún indiferenciadas. La corteza cerebelosa alcanza sus dimensiones definitivas después del nacimiento. Los núcleos dentados y dentados accesorios (emboliforme, globoso y fastigio) se presencian antes del nacimiento. Posteriormente, los axones que salen de estos núcleos cruzan el mesencéfalo para llegar al prosencéfalo y constituyen el pedúnculo cerebeloso superior. El crecimiento axonal de las fibras corticopontinas y pontocerebelosas que conectan las cortezas cerebral y cerebelosa conlleva la formación del pedúnculo cerebeloso medio. Axones sensitivos provenientes de la médula espinal, núcleos olivares y vestibulares forman el pedúnculo cerebeloso inferior.
MESENCEFALO
El mesencéfalo constituye la vesícula encefálica inmediatamente cefálica al rombencéfalo que sufre menos modificaciones durante el desarrollo del SNC. La cavidad de la vesícula mesencefálica se reduce considerablemente para formar un conducto que unirá los futuros III y IV ventrículos: el acueducto cerebral (de Silvio).
A cada lado, las placas basales y alares están separadas por el surco limitante. Cada placa basal tiene 2 grupos de motoneuronas: (1) Eferente somático o medial: origina los nervios craneales III y IV (2) Eferente visceral general: forma el núcleo de Edinger-Westphal. La capa marginal de las placas basales se expande y origina el pie de los pedúnculos cerebrales por donde descienden fibras desde la corteza cerebral a centros motores inferiores del puente y médula espinal (tractos corticopontinos, corticobulbares y corticoespinales). Las placas alares y del techo forman el tectum. Neuroblastos de las placas alares migran a la capa marginal del tectum y forman agregados estratificados de neuronas sensitivas separadas por un surco transverso: los colículos anteriores (superiores) y los colículos posteriores (inferiores). Estos últimos son centros de relevo para reflejos auditivos, mientras que los colículos anteriores forman centros de correlación y de reflejos para estímulos visuales.
Una banda de sustancia gris adyacente al pedúnculo cerebral forma la sustancia negra (locus niger). Se tienen muchas dudas si este elemento se origina a partir de células de las placas basales o de células de las placas alares que migran ventralmente.
PROSENCEFALO
Antes del cierre del neuroporo craneal, aparecen dos divertículos laterales a cada lado del prosencéfalo: las vesículas ópticas (primordios de la retina y nervios ópticos). Posteriormente, aparecen las vesículas telencefálicas que formarán los hemisferios cerebrales y sus cavidades: los ventrículos laterales. Las placas del piso y del techo son delgadas; por otra parte, las paredes laterales son gruesas como en la médula espinal.
DIENCEFALO
Se desarrolla a partir de la porción media del prosencéfalo y consta de placas del techo y alares pero carece de placas basales y del piso. El mesénquima vascularizado de los ependimocitos de la placa del techo origina el plexo coroideo del tercer ventrículo. La porción caudal de la placa del techo forma un divertículo ubicado anteriormente al mesencéfalo que hacia la séptima semana ya forma un órgano macizo con forma de cono: el cuerpo pineal (epífisis). El calcio acumulado en la epífisis durante la adultez la hace un importante punto de referencia en imagenología. Tiene importantes funciones sexuales (secreción de melatonina que estimula la secreción de gonadotropinas) y es posible que forme un nexus entre los ciclos de luz solar y los ciclos endocrinos y de la conducta.
En las paredes laterales del Tercer ventrículo (placas alares del diencéfalo) aparecen tres prominencias que posteriormente formarán el hipotálamo tálamo y epitálamo. El surco hipotalámico divide las placas alares en una porción ventral (hipotálamo) y una dorsal (tálamo). Este surco no es la continuación del surco limitante y no divide porciones sensitivas y motoras.
La notable proliferación ocurrida en el tálamo hace que éste protruya hacia el III ventrículo de modo que las regiones talámicas derecha e izquierda se fusionan en la línea media formando la adhesión intertalámica (presente en un 70 a 80% de los cerebros).
El hipotálamo (porción inferior de la placa alar) se diferencia en varios grupos nucleares que constituyen centros reguladores de variadas funciones viscerales (temperatura corporal, emociones, digestión, sueño, etc.). Uno de estos núcleos, los cuerpos mamilares, sobresalen en la superficie ventral del hipotálamo a cada lado de la línea media.
El techo y la parte dorsal de la pared lateral del diencéfalo formarán el epitálamo.
Desarrollo de la Hipófisis (glándula pituitaria): La hipófisis se origina totalmente del ectodermo (cuarta semana). Se desarrolla a partir de dos porciones: (1) Una evaginación diencefálica hacia caudal (Infundíbulo). (2) Una evaginación ectodérmica del estomodeo (cavidad bucal primitiva) anterior a la membrana bucofaríngea (Bolsa de Rathke). Este doble origen explica la diferencia de tejidos hipofisiarios. En la tercera semana, la bolsa de Rathke crece dorsalmente hacia el infundíbulo. Al final del segundo mes, pierde contacto con la cavidad bucal y se contacta íntimamente con el infundíbulo. Posteriormente, la multiplicación de las células de la pared anterior de la bolsa de Rathke originan el lóbulo anterior de la hipófisis (Adenohipófisis). La adenohipófisis consta de tres partes: (1) Una prolongación de este lóbulo que crece rodeando el tallo del infundíbulo: la pars tuberalis. (2) la pars distalis que constituye el lóbulo anterior propiamente tal. (3) la pared posterior de la bolsa de Rathke no prolifera y forma la pars intermedia, de poca importancia en el humano.
La evaginación diencefálica origina la eminencia media, el tallo infundibular y la pars nerviosa que en conjunto se les denomina Neurohipófisis (lóbulo posterior). El extremo distal del neuroepitelio del infundíbulo experimenta una proliferación que origina los pituicitos, las células de sostén de la neurohipófisis.
La proliferación de la pared anterior de la bolsa de Rathke reduce su luz hasta formar una pequeña hendidura (ella es un posible sitio de quistes, por ello, es importante conocerla).
TELENCEFALO
Es la vesícula encefálica más rostral. Consta en 2 evaginaciones laterales (hemisferios cerebrales) y una porción media (lámina terminal). Sus cavidades (ventrículos laterales) comunican con el III ventrículo a través de los agujeros interventriculares.
Los Hemisferios Cerebrales: Entre la 5º y12º semana, las evaginaciones bilaterales de la pared lateral del prosencéfalo originan los hemisferios cerebrales. La expansión anterior forma los lóbulos frontales mientras la superolateral origina los lóbulos parietales; finalmente, la expansión posteroinferior forma los lóbulos temporales y occipitales. El proceso continúa con un aplanamiento medial de los hemisferios cerebrales.
El mesénquima de la fisura longitudinal del cerebro origina la hoz del cerebro (falx cerebri). La pared medial de los hemisferios -donde se unen al techo diencefálico- es delgada y sólo consta de una capa de células ependimarias cubierta de mesénquima vascularizado: el plexo coroideo del ventrículo lateral.
Durante la 6º semana, la parte basal de los hemisferios aumenta de tamaño y sobresale hacia el ventrículo lateral dando origen al Cuerpo estriado. Esta región de la pared hemisférica se expande en dirección posterior y se divide en 2 partes: (1) núcleo caudado, dorsomedialmente (2) núcleo lenticular, ventrolateralmente. La fusión de la pared medial del hemisferio y la pared lateral del diencéfalo permite el contacto entre el núcleo caudado y tálamo. Tractos ascendentes y descendentes de la corteza cerebral pasan entre tálamo y núcleo caudado medialmente y núcleo lentiforme lateralmente formando la cápsula interna. La pared del prosencéfalo se engrosa formando una estructura longitudinal que protruye al ventrículo lateral: el hipocampo.
La zona suprayacente al núcleo lentiforme crece lentamente y queda ocultada entre los lóbulos temporal y occipital (isla o ínsula de Reil). Al final de la vida fetal, la superficie hemisférica crece tan rápido que se forman giros (circunvoluciones) separados por surcos y cisuras. Estos surcos y giros permiten un aumento considerable de la superficie cerebral -por ende, un aumento de la superficie cortical- sin sobrepasar el volumen del cráneo.
MEDULA ESPINAL:
Configuracion Y Divisiones
La médula espinal es la parte del SNC que se aloja en el canal raquídeo o vertebral desde el foramen magnum hasta el borde superior del cuerpo de la segunda vértebra lumbar (L2).
Tiene forma cilíndrica y su aspecto externo es blanquecino debido a que superficialmente está compuesta de fibras nerviosas mielinizadas. Su longitud alcanza un promedio de 45 cm. Su ancho va cambiando según la cantidad de fibras que lleven sus tractos. De este modo, se observa que a nivel cervical, precisamente en donde se originan las raíces que constituyen el plexo braquial, la médula espinal se encuentra notablemente aplanada en sentido anteroposterior formando un engrosamiento fusiforme, el mayor de la médula espinal: el engrosamiento cervical (C3-T2). Asimismo, a nivel lumbar donde se origina el plexo lumbosacro, la médula espinal presenta el engrosamiento lumbar (L1-S3). El extremo inferior de la médula termina en forma de cono: el cono medular. Una fina banda de tejido fibroso, el filum terminale, avanza en medio de la cauda equina hasta unirse al periostio del dorso del cóccix. El filum terminale es una prolongación de la piamadre que indica el camino de regresión de la médula espinal y tiene la función de fijar el extremo inferior de la médula espinal. Bajo el cono medular y hasta la terminación del saco dural en el borde inferior de S2, el espacio subaracnoideo sólo contiene la cauda equina y el filum terminale flotando en líquido cefalorraquídeo (LCR). Por tal razón, esta es la zona con menor riesgo para efectuar una punción lumbar. La cauda equina la forman las raíces dorsales y ventrales correspondientes a los segmentos lumbares, sacro y coccígeo que corren verticalmente bajo el cono medular.
Al desprender las meninges que cubren la médula espinal, se observa en ella un tenue surco que recorre la línea media posterior (surco mediano posterior) y una fisura profunda que va por la línea media anterior (fisura mediana anterior). Ambos elementos dividen a la médula en dos mitades relativamente simétricas. Los 31 pares de nervios espinales o raquídeos se unen a la médula espinal a través de sus raíces posteriores (sensitivas) y anteriores (motoras). Cada raíz consta de un grupo de raicillas que se van expandiendo a medida que se acercan a la médula, llegando a abarcar la totalidad de su segmento medular. El lugar donde las raicillas posteriores penetran a la médula espinal está indicado por un surco poco profundo, el surco lateral posterior. Por otra parte, las raicillas anteriores no emergen por un lugar específico indicado por un surco, aunque algunos autores sí hacen referencia al surco lateral anterior. Con propósitos descriptivos, se ha dividido cada mitad de la médula espinal en tres columnas de sustancia blanca (columnas blancas, cordones o funículos) tomando como límites los diferentes surcos y fisuras: (1) Cordón posterior, entre el surco mediano posterior y el surco lateral posterior (2) Cordón lateral, entre el surco lateral posterior y la salida de las raicillas anteriores de los nervios espinales (3) Cordón anterior, entre la salida de las raicillas anteriores y la fisura mediana anterior. En los segmentos medulares superiores cada cordón posterior está dividido por el tabique intermedio posterior que en la superficie está indicado por el tenue surco intermedio posterior.
Debido al crecimiento desproporcionado de la médula espinal y la columna vertebral, la médula sólo alcanza el borde superior del cuerpo de L2 en su extremo caudal. Es sabido que durante la 12º semana de vida intra uterina la médula espinal abarca todo el canal vertebral, pero debido al acelerado crecimiento vertebral, el extremo inferior de la médula progresivamente va alcanzando niveles máscefálicos. Al momento del nacimiento la médula se encuentra a nivel de L3, pero como el proceso de crecimiento vertebral continúa unos años, finalmente la médula alcanza el borde superior de L2 en los individuos adultos. Consecuencia de este crecimiento diferencial es que los segmentos medulares no se corresponden con los segmentos vertebrales: a los segmentos vertebrales desde C1 a C3 le corresponden los segmentos medulares C1 a C3; a los segmentos vertebrales C4 a C7 se les suma 1 y se obtiene el segmento medular ; por ejemplo, ala vértebra C4 está frente al segmento medular C5; a las vértebras T1 a T6 se les agrega 2 para obtener el segmento medular correspondiente ; por ejemplo, a la vértebra T1 le corresponde el segmento medular T3; a las vértebras desde T7 a T9 se les debe sumar 3 para tener el segmento medular ; por ejemplo, a la vértebra T7 le corresponde el segmento medular T10; a las vértebras T10 a T12 les corresponden los segmentos medulares desde L1 hasta L5; finalmente, a las vértebras L1 y borde superior de L2 les corresponde el segmento medular sacro-coccígeo. Por otra parte, los nervios espinales de cada segmento medular siempre salen por el respectivo agujero intervertebral. Si bien las raíces de los nervios cervicales corren horizontalmente, consecuencia del crecimiento diferencial vertebromedular las raíces de los nervios espinales hacen su recorrido cada vez más vertical mientras más bajo es el segmento medular.
Estructura Interna De Médula Espinal:
En un corte transversal se observa que la médula espinal consta de una región central con forma de H llamada sustancia gris y una región periférica de aspecto blanquecino, la sustancia blanca. Las prolongaciones posteriores de la sustancia gris son relativamente delgadas y alcanzan practicamente el surco lateral posterior, ellas son las astas posteriores. Las prolongaciones anteriores anchas y redondeadas se denominan astas anteriores.
Astas anteriores y posteriores
La disposición tridimensional de las astas anteriores y posteriores conforma verdaderas columnas que recorren la médula espinal para constituir las columnas grises anterior y posterior. Las astas posteriores, funcionalmente somatosensitivas, están formadas por neuronas sensitivas que reciben los impulsos que llegan por las raíces posteriores. Las astas anteriores, funcionalmente somatomotoras, están constituidas por neuronas motoras cuyos axones salen por las raíces anteriores. En los segmentos torácicos y lumbares superiores existe una pequeña asta lateral que emerge de la unión del asta anterior con la posterior y contiene neuronas viscerales simpáticas. En la parte lateral de la base del asta posterior de los segmentos cervicales superiores es difícil distinguir la sustancia gris de la blanca debido a que células y fibras nerviosas de varios tipos se encuentran mezcladas: es la formación reticular que se continúa superiormente como la formación reticular del tronco encefálico.
Las columnas grises anterior y posterior de cada lado se encuentran unidas por una banda transversal de sustancia gris: la comisura gris. Justo en medio de la comisura gris se encuentra un pequeño conducto lleno de LCR que recorre completamente a la médula espinal: el canal central (también llamado canal del epéndimo). Este conducto suele ser apenas visible o permanecer ocluido con el epitelio cilíndrico ciliado que lo recubre (epéndimo). Superiormente, se continúa con el canal central de la mitad caudal del bulbo raquídeo para luego abrirse paso a la cavidad del IV ventrículo. En el extremo inferior de la médula espinal puede formar una dilatación que se prolonga hacia el filum terminale (ventrículo terminal). El canal central sirve como referencia para dividir la comisura gris en dos mitades en sentido anteroposterior: la comisura gris anterior y la comisura gris posterior. Entre la comisura gris anterior y la fisura mediana anterior existe una banda transversal de sustancia blanca que comunica ambos cordones anteriores, la comisura blanca anterior. Los cordones posteriores están totalmente separados por el tabique mediano posterior que va desde el surco mediano posterior hasta la comisura gris posterior.
La cantidad de sustancia gris presente en los diferentes niveles medulares depende de la cantidad de músculos inervados por ese nivel. Por tal motivo, en los segmentos torácicos las astas son delgadas y pequeñas, en cambio, en los segmentos cervicales inferiores y lumbosacros son más gruesas ya que contienen las neuronas motoras y sensitivas asociadas a los plexos braquial y lumbosacro. En estos segmentos, los vértices de las astas posteriores presentan un área de sustancia gris con apariencia gelatinosa (sustancia gelatinosa) que también está en los segmentos torácicos, pero allí es difícil observarla debido a su escasez.
Tipos De Neuronas
La sustancia gris de la médula espinal está formada por un conjunto de somas neuronales multipolares, células gliales y vasos sanguíneos. Las neuroglias se encargan de formar una intrincada red que envuelve, nutre y soporta a los elementos neuronales. Existe un número considerable de agrupaciones celulares bien definidas:
(1) Núcleos de las astas anteriores: La mayoría de las neuronas de las columnas grises anteriores son multipolares con grandes prolongaciones y abundante sustancia de Nissl. Sus axones eferentes alfa forman las raíces anteriores de los nervios espinales e inervan a los músculos esqueléticos. Por otra parte, los axones eferentes gamma son las prolongaciones de las neuronas multipolares más pequeñas del asta anterior que van a inervar las fibras intrafusales de los husos musculares.
Los grupos celulares más mediales de la columna gris anterior inervan la musculatura axial, mientras que los grupos laterales inervan las extremidades; esto explica la prominencia en la parte lateral del asta anterior en los segmentos que originan los plexos braquial y lumbosacro. Los grupos celulares relativos a los miembros que inervan los músculos proximales están dispuestos medialmente, mientras los que inervan a los músculos distales están lateralmente. Por esto mismo, es posible que algunas lesiones produzcan parálisis de un grupo muscular sin afectar a otro.
Existen descripciones citoarquitectónicas que nombran una infinidad de grupos columnares, pero esto no tiene mucha aplicación clínica. Por tal motivo, es más práctico dividir la columna gris anterior en tres grupos: (a) el grupo lateral se encuentra en los segmentos medulares cervicales y lumbosacros e inerva a la musculatura de las extremidades. (b) el grupo central es el más pequeño y se encuentra en algunos segmentos cervicales y en los lumbosacros. En los segmentos medulares C3, C4 y C5 es posible observar el núcleo frénico que da inervación al diafragma. En los cinco o seis segmentos cervicales superiores se forma el núcleo accesorio el cual proporciona la inervación a los músculos esternocleidomastoideo y trapecio. Los axones de las neuronas que forman el núcleo accesorio originan la raíz espinal del nervio accesorio (XI).(c) el grupo medial se encuentra en la mayoría de los segmentos medulares. Se encarga de inervar los músculos del cuello y del tronco, incluyendo los músculos intercostales y abdominales.
(2) Núcleos de las astas posteriores: (a) la sustancia gelatinosa está formada por pequeñas neuronas Golgi tipo II distribuidas en una red neuroniana. Recibe estímulos exteroceptivos que vienen por las raíces posteriores, entre ellos, de dolor y temperatura (estímulos termalgésicos). Se ubica en los vértices de las astas posteriores a lo largo de la médula espinal, pero en los dos primeros segmentos cervicales es un componente notable que recibe impulsos de la raíz sensitiva del nervio trigémino y se le denomina núcleo espinal del V par. (b) Anterior a la sustancia gelatinosa se ubica un importante grupo celular que se encuentra a lo largo de toda la médula espinal, se trata del núcleo propio el cual recibe estímulos propioceptivos y tactiles y tambien cumple funciones de asociación intersegmentaria. (c) En el centro del asta posterior se ubican algunas interneuronas y unas pequeñas neuronas receptoras de estímulos exteroceptivos. (d) En la porción medial de la base del asta posterior desde el octavo segmento cervical hasta el tercer o cuarto segmento lumbar está el núcleo torácico, también denominado nucleus dorsalis o columna de Clark. Este núcleo recibe estímulos propioceptivos desde los husos musculares y tendinosos que van al cerebelo por los tractos espinocerebelosos. (e) Los núcleos viscerales aferentes están formados por pequeñas neuronas que se encuentran ubicadas lateralmente al núcleo torácico desde los segmentos torácicos al tercer segmento lumbar (asta lateral). Se relacionan con la recepción de información visceral aferente que llega por las raíces posteriores. Las neuronas visceromotoras del asta lateral envian sus axones acompañando a las fibras de las neuronas somatomotoras constituyendo ambas la raíces anteriores de los nervios raquídeos.
Las Meninges De La Médula Espinal
La médula espinal, al igual que el encéfalo, está envuelta por las meninges: duramadre, aracnoides y piamadre.
La duramadre es una membrana fuerte, densa y fibrosa que envuelve la médula espinal y la cauda equina. Se continúa a través del foramen magno con la capa de duramadre que envuelve al encéfalo. La duramadre se encuentra separado laxamente de las paredes del canal vertebral por el espacio epidural (también llamado espacio extradural o peridural). Este espacio es ocupado por tejido conjuntivo laxo y por un plexo venoso. Los ganglios de la raíz posterior se ubican dentro del agujero intervertebral, y justo por fuera de él se unen ambas raíces para formar un nervio raquídeo mixto. Cuando ambas raíces atraviesan la duramadre y la aracnoides a nivel del agujero intervertebral, la duramadre se adhiere fuertemente a cada nervio formando una capa continua con el epineuro.
La aracnoides es una membrana delgada e impermeable que recubre totalmente la médula espinal. Se ubica entre la piamadre, más profunda, y la duramadre, más superficial. Entre la piamadre y la aracnoides existe un espacio bastante amplio: el espacio subaracnoideo. Este espacio rodea a todo el encéfalo y prosigue inferiormente a través del foramen magno hasta el borde inferior de S2, donde la duramadre y la aracnoides se fusionan con el filum terminale no dejando espacio alguno. Su importancia radica en que contiene el Líquido Cefalorraquídeo o Cerebroespinal. El espacio subaracnoídeo es atravesado por finas trabéculas aracnoideas que se unen a la piamadre. Como la aracnoides se continúa con las raíces de los nervios espinales hasta su salida por el agujero intervertebral, se forman unas pequeñas extensiones laterales del espacio subaracnoideo.
La piamadre es una capa única y delgada de carácter vascular que se adosa íntimamente a la médula espinal. Por las caras laterales de la médula, a igual distancia entre las raíces dorsales y ventrales de los nervios espinales, unas 22 extensiones membranosas puntiformes de la piamadre van a insertarse firmemente a la cara interna de la duramadre y de la aracnoides: son los ligamentos dentados. Ellos facilitan la suspensión de la médula espinal justo en medio del saco dural. En tal labor también colaboran (1) la continuidad con el tronco encefálico (2) la presión ejercida por el LCR (3) el filum terminale.
Irrigación y Drenaje De Médula Espinal:
El patrón básico de irrigación arterial de la médula espinal consiste en tres vasos que corren longitudinalmente a lo largo de ella extendiéndose hasta el cono medular y cuyo origen es en el extremo cefálico de la médula cervical. Sin embargo este sistema longitudinal de vasos es reforzado por vasos tributarios transversales que penetran al canal medular; a través de los forámenes intervertebrales junto a los nervios y raíces espinales posteriores y anteriores.
Las anastomosis entre los vasos longitudinales (arteria espinal anterior o arterias espinales posteriores) y los vasos segmentarios se produce en la superficie de la médula espinal.
De los 31 pares de vasos segmentarios que penetran con los nervios raquídeos no todos terminan en la médula espinal. De todos ellos se pueden distinguir:
1. Arterias radiculares propiamente tales: es decir, arterias que rrigan las raíces nerviosas y el ganglio sensitivo solamente, por lo tanto no alcanzan la médula.
2. Arterias radículo piales: Son aquellas que llegan sólo hasta la piamadre.
3. Arterias radículo medulares: Son las únicas que llegan a irrigar a la médula, anastomosándose con el sistema longitudinal. Ellas son relativamente escasas, en un número de 8 a 10. Ingresan, por lo general, por un sólo lado de la médula ya sea el derecho o el izquierdo y frecuentemente se dividen en dos ramas, una anterior y otra posterior, las que acompañan a las respectivas raíces nerviosas.
En la médula espinal se distinguen tres territorios o segmentos de acuerdo a su circulación: superior o cérvico-toráxico, intermedio o toráxico medio e inferior o tóraco lumbar
A. El segmento superior, comprende toda la médula cervical hasta el segundo o tercer segmento toráxico.
Desde C1 hasta C4 la médula espinal es irrigada por la arteria espinal anterior, que deriva de la fusión de dos arterias espinales ramas de las vertebrales. Ademas tenemos dos arterias espinales posteriores que también provienen de las arteria vertebrales. La porción media del segmento superior es irrigada por 2 ó 3 arterias segmentarias que nacen directamente de las vertebrales. La porción más inferior del segmento superior es irrigada por una rama de la arteria cervical profunda. Para asegurar la circulación de este territorio se producen una serie de anastomosis entre distintos vasos del cuello principalmente a través de la arteria cervical profunda rama del tronco costocervical y arteria cervical ascendente rama de la tiroídea inferior la cual se origina en el tronco tirocervical. Por lo tanto, frente a una obstrucción en la región del engrosamiento cervical el déficit puede ser suplido por alguna de las numerosas colaterales antes mencionadas.
B. El territorio intermedio de la médula espinal se extiende entre los segmentos T4 y T8.
Generalmente hay sólo una rama del sistema segmentario, a nivel de T7 aproximadamente,la cual proviene de una arteria intercostal rama de la aorta. Este territorio es el más pobremente irrigado por lo tanto, es el más lábil de los tres segmentos mencionados, frente a cualquier obstrucción vascular.
C. Por último, el territorio medular inferior que va desde los últimos segmentos toráxicos hasta el cono medular, depende en gran parte de una arteria radículo medular llamada arteria radicular mayor o de Adamkiewicz que es la de mayor diámetro de todas las arterias radiculo-medulares. Es rama de las primeras lumbares, prveniente de la aorta, llega a la médula con mayor frecuencia por una de las raíces del lado izquierdo entre los segmentos T12 y L4 irrigando el engrosamiento lumbar. A nivel del cono medular se forma una amplia red anastomótica llamada asa del cono medular, formada por 1 ó 2 arterias que viajan con alguna de las raíces de la cola de caballo y que vienen de ramas de las sacrales media y lateral, para unirse a las tres arterias espinales. Como se puede ver, éste es un territorio con muy buena vascularización.
Sistema arterial intramedular.
El sistema arterial intramedular está formado por las arterias centrales y por el plexo perimedular
Las arterias centrales nacen en ángulo recto de la espinal anterior, se dirigen por la fisura medial anterior hasta la comisura blanca anterior, lugar donde se desvían a uno u otro lado. Son varias por centímetro, siendo menos numerosos y de menor calibre en el territorio medular intermedio (toráxico medio).Irrigan la parte más profunda de la sustancia blanca y la sustancia gris menos las puntas de las astas posteriores. Son funcionalmente arterias terminales.
El plexo perimedular está formado por varias arteriolas anastomóticas provenientes de espinales posteriores, anterior y radículo piales. Desde este plexo existen numerosos vasos que penetran en forma radial a la médula, constituyendo la corona radiada.
Las arterias espinales posteriores y sus ramos penetrantes irrigan las puntas de las astas posteriores y cordón posterior.Estas últimas al igual que las arterias centrales son vasos funcionalmente terminales, sin embargo, hay una zona más o menos circular a la médula en que se pueden superponer ambos sistemas.
Sistema Venoso
A lo largo de la columna vertebral se extienden dos grandes plexos: los plexos vertebrales interno y externo, que forman anillos definidos en torno a cada nivel vertebral. Existe libre comunicación entre ambos. Algunas ramas provenientes de las vértebras, ligamentos y médula espinal llegan a conformar el plexo. Son importantes las variaciones de presión del LCR o de la cavidad torácica, ya que estas venas son relativamente pobres en válvulas y son posibles grandes variaciones en el volumen sanguíneo de ellas.
Plexo vertebral interno: Es una red anastomótica de venas que se entremezcla con el tejido conjuntivo laxo del espacio epidural. Se observan venas que corren longitudinalmente por la superficie de los cuerpos vertebrales y discos intervertebrales a cada lado del ligamento longitudinal posterior. Entre este ligamento y el cuerpo vertebral existen ramas que reciben la sangre proveniente del terrritorio de las arterias basilar y vertebrales.
La configuración del sistema venoso medular es muy similar al arterial. Sin embargo, se diferencian en que las venas centrales son menos numerosas que las arterias y drenan un territorio más reducido (la parte más anterior de la sustancia gris y de la sustancia blanca). Las venas penetrantes, por lo tanto, drenan la mayor parte de la sustancia gris y blanca y terminan en el plexo perimedular que conecta a las venas espinales anterior y posteriores.
La vena espinal anterior, más superficial que la arteria, recibe la sangre del territorio de las arterias centrales y de la porción anterior del plexo perimedular. Se vacia en las venas radiculares anteriores que se encuentran cada dos o tres segmentos.
Por último, las venas radiculares anterior y posterior se anastomosan con las venas de los plexos vertebrales y conforman las venas intervertebrales. Estas venas drenan la mayor parte de la sangre de la médula espinal y de los plexos vertebrales hacia las venas vertebrales, intercostales, lumbares y sacra lateral, dependiendo de qué territorio se trate.
3. VIAS ASCENDENTES MEDULARES, GANGLIO ESPINAL Y TIPOS DE SENSIBILIDAD:
Ganglio espinal
El ganglio nervioso es el conjunto de neuronas localizadas fuera de los órganos del sistema nervioso central. El ganglio espinal, localizado en el trayecto de la raíz posterior, está formado por neuronas monopolares o pseudo-monopolares cuya fibra, luego de su nacimiento da una serie complicada de tortuosidades que en conjunto reciben el nombre de glomérulo. La fibra se divide luego en dos ramas divergentes, una lateral, que unida a la raíz anterior forma el nervio raquídeo, y una medial, que con sus similares forma la raíz posterior y penetra a la médula, a través del surco lateral dorsal. El cuerpo de las células ganglionares está rodeado por una membrana o cápsula de células aplanadas que se continúa con el neurilema de la fibra, dentro de la cápsula, y al lado de la neurona. Existen algunas células pequeñas de función desconocida y llamadas células satélites.
La ramificación perisférica de la neurona sensitiva termina en los distintos receptores cutáneos para captar las condiciones exteriores, o en los músculos y articulaciones, y reconocer la posición de nuestro cuerpo y sus segmentos, o en las vísceras para regular su estado de funcionamiento de acuerdo con las circunstancias de cada momento.
Supongamos ahora que por descuido nos pinchamos con una espina el pie. Enseguida lo retiramos por el dolor. El pinchazo ya ha sido recibido por un receptor de dolor de la superficie cutánea e inmediatamente se ha convertido en impulso. Los impulsos van desde el receptor, a través de largas terminaciones de células, hasta su centro, el ganglio espinal, al lado de la columna vertebral; de allí prosigue a la médula. Las células nerviosas se extienden desde la punta de los dedos hasta la columna vertebral. Cuando el impulso llega a la médula espinal, esta reacciona con un reflejo que ordena la retirada del dedo. La respuesta refleja no sigue el mismo camino que el impulso nervioso, pues las fibras nerviosas suelen ser de una sola dirección. El cerebro aun no ha recibido la sensación de dolor cuando el acto reflejo se ha producido. Hará falta una fracción de segundo para que la médula le informe y el llegue a la reflexión: "me duele, luego, he debido pincharme". El cerebro reconoce la sensación gracias a su memoria almacenada por acontecimientos y sensaciones anteriores. Los reflejos son mas rápidos, vitales y primitivos que la reflexión, y en los animales terrestres juegan un papel; importantísimo. Cuando tragamos saliva, cerramos un ojo para evitar que nos entre una brizna o estornudamos, porque se han irritado los receptores de la nariz, estamos actuando por reflejos. Quizá muchas de las cosas que hacemos sean una desconocida reacción ante estímulos externos. Sin saberlo, tenemos frecuentes reflejos inconscientes; la sed y el hambre, la alegría y la tristeza no sufren ningún proceso de control.
Exterocepción:
Se relaciona con las sensaciones de calor, frío, dolor y forma gruesa de sensación táctil que llegan a las astas posteriores. La sustancia gelatinosa se relaciona en parte con la recepción de impulsos dolorosos a través de fibras amielínicas y poco mielinizadas que constituyen el tracto posterolateral (de Lissauer) ubicado entre el vértice del asta posterior y la superficie de la médula espinal cercano a las raíces posteriores. Todas las células de la columna gris posterior que reciben impulsos exteroceptivos originan axones que se decusan pasando por la comisura blanca anterior al lado opuesto, en donde se separan en dos tractos ascendentes: Los tractos espinotalámicos lateral y anterior.Fig.4
Propiocepción:
La propiocepción se produce por estimulación de los diversos receptores ubicados en los músculos, tendones y articulaciones. Los impulsos aferentes que van al cerebelo para la coordinación inconsciente suben por los tractos espinocerebelosos (cordón lateral de la sustancia blanca). Los que van a la corteza cerebral intervienen en el control cortical de los movimientos originando impresiones conscientes. Estos ascienden por el cordón posterior de la médula espinal (fascículos gracilis y cuneatus).Fig.3
Viscerocepción Y Vías Viscerales:
El dolor visceral es un tipo de dolor periférico proveniente de las vísceras toracoabdominales que se localiza sobre los tejidos que recubren el órgano que lo origina, en especial sobre la piel del tórax y del abdomen, o a una distancia próxima al sitio donde se produce. Esto último es lo que se designa como dolor referido o heterotópico, ya que se refiere de una víscera que lo produce a una región parietal próxima en conexión fisiológica con ella. En realidad, el dolor referirá al dermatoma que originó el órgano en el desarrollo embrionario (regla de los dermatomas). Por ejemplo, el dolor cardíaco es referido a la porción anterior del brazo izquierdo, lo que se explica por el hecho que el corazón y el brazo tienen el mismo origen segmentario. El dolor proveniente de las vísceras toracoabdominales se ha constituído en un elemento importante en el establecimiento del diagnóstico de las patologías que afectan a estos órganos. A diferencia del dolor superficial, el dolor visceral provocado por lesiones muy localizadas rara vez es intenso, pero si se provoca una estimulación difusa de los receptores de dolor en una extensa área de la víscera el dolor puede ser muy intenso como ocurre en las isquemias, por ejemplo. Los dolores viscerales suelen ser causados por distensión excesiva, espasmos de la musculatura lisa de una víscera hueca, daño químico o isquemia y son transmitidos hasta la médula espinal por neuronas cuyos somas se encuentran ubicados en un ganglio de la raíz posterior. La dendrita de estas neuronas contacta en la víscera con receptores de dolor y estiramiento; por otra parte, el axón penetra por la raíz posterior y sinapta con neuronas del asta posterior o lateral. Las fibras provenientes de estas neuronas ascienden junto a los tractos espinotalámicos , posiblemente através del tracto espinoreticular hasta contactar con nucleos intralaminares del tálamo, de aquí los axones se dirigen a la corteza cerebral.
Se ha sugerido que el dolor visceral referido se produce de la siguiente manera: Las fibras de tipo C que llevan dolor visceral a un determinado segmento de la médula espinal junto a las fibras que llevan dolor desde la piel pueden sinaptar en una misma neurona de segundo orden o en neuronas vecinas de las astas posteriores. Por lo tanto, es posible que algunas de las sensaciones viscerales dolorosas sean conducidas por algunas de estas neuronas en común para ambas vías y que el paciente sienta como si el dolor de la víscera se estuviese produciendo en una determinada región de la piel.
Receptores Sensitivos Y Tipos De Sensibilidad:
El reflejo es la unidad fisiológica del sistema nervioso. Se define como una respuesta motriz de tipo involuntaria que ocurre inmediatamente después de aplicar un estímulo en particular, y que puede ser o no consciente. Si la respuesta no es inmediata no puede ser considerada un reflejo. Otra característica esencial de la respuesta refleja es que parece presentarse y ejecutarse con un fin determinado, y la respuesta se coordina y adapta en vista de tal fin. Su realización es posible gracias al arco reflejo, cuyos componentes básicos son: (1) un receptor (2) una neurona aferente (3) una neurona eferente (4) un órgano efector. Cuando este arco reflejo sólo involucra una sinapsis, se denomina arco reflejo monosináptico; por ello, el tiempo entre la aplicación del estímulo y la ejecución del reflejo es muy pequeño (período latente breve), como sucede en el reflejo patelar. Los arcos reflejos se encargan de la mantención del tono muscular y es por ello que permiten una postura corporal adecuada.
Las fibras de las raíces posteriores que llegan hasta el asta posterior presentan diferente grosor según el estímulo que ellas transmitan. Las fibras que transmiten aferencias exteroceptivas son de pequeño calibre y poco mielínicas, mientras las que llevan impulsos propioceptivos son más gruesas y mielinizadas. Las neuronas que sinaptan con la sustancia gelatinosa son amielínicas. Las fibras de la raíz posterior toman contacto con las motoneuronas del asta anterior directa o indirectamente a través de interneuronas que tienen sus somas en el asta posterior. Los axones de las interneuronas cruzan por la comisura blanca anterior para conectarse con las motoneuronas del lado opuesto (base de los reflejos segmentarios cruzados).
Una característica importante de los reflejos es su localización perfecta y estricta. Cada reflejo origina una respuesta que ocurre siempre en el mismo sitio. Se ha demostrado que luego de la descarga normal de la neurona eferente sobre el órgano efector viene un periodo prolongado de descarga asincrónica. Este suceso se explica por la presencia de colaterales del axón de la neurona aferente que sinaptan con interneuronas que vuelven a sinaptar con la neurona eferente produciendo una descarga prolongada luego del impulso inicial. Luego de ocurrido un reflejo viene un período refractario en el cual no es posible una respuesta refleja ante un estímulo. Otra característica de los reflejos es que se fatigan fácilmente. Los centros reflejos son muy susceptibles a la anoxia y a ciertos fármacos; es por ello que la ausencia o cualquier alteración de la actividad refleja juega un papel preponderante en el diagnóstico clínico.
Existen dos propiedades de los reflejos espinales que deben considerarse: (1) la ley de la inervación recíproca indica que los reflejos extensor y flexor de un mismo miembro no pueden realizarse simultáneamente. Se cree que la neurona aferente que llega al músculo flexor envía una cierta cantidad de colaterales al músculo extensor para inhibirlo. (2) el reflejo de extensión cruzado: al provocar el reflejo de flexión en el miembro inferior de un lado, el miembro del otro lado se hiperextiende. Por ejemplo, si se estimula alternativamente la planta de un pie y del otro, se produce un movimiento de pedaleo. Estas respuestas también han sido observadas en el miembro superior, pero son menos frecuentes.
Los reflejos segmentarios son fuertemente influenciados por centros neuronales superiores a través de los tractos descendentes largos (ver más adelante). Por ejemplo, la sección transversal de la médula espinal involucra la pérdida de estas influencias y causa un estado de depresión funcional de toda la región corporal que depende de los segmentos medulares que se ubican por debajo de la sección, caracterizada por parálisis flácida total, gran vasodilatación con hipotensión arterial, incontinencia urinaria y fecal y ausencia de reflejos. Una vez pasada esta etapa de shock espinal, vuelven los reflejos segmentarios y el tono muscular se ve aumentado. Esta rigidez de decerebración es producida por una hiperactividad de las fibras eferentes gamma sobre los husos musculares, descontroladas porque no tienen control de los centros superiores (automatismo medular). Nos hemos referido a un tema bastante controvertido: Se admite corrientemente que la acción de la corteza motora y la vía piramidal inhiben el tono muscular. Es por ello que en la rigidez de decerebración por lesión de la vía corticoespinal se observa una marcada hipertonía.
Fibras de las raíces posteriores no sólo terminan en su segmento medular. Una considerable parte de ellas se bifurca al penetrar a la médula y se divide en ramas ascendentes y descendentes que emiten colaterales hacia el asta posterior hasta que ellas mismas terminan en la sustancia gris de segmentos arriba o abajo. Muchas ramas descendentes se agrupan formando tractos que corren en la columna blanca posterior y establecen conexiones intersegmentarias. Las interneuronas del asta posterior extienden sus axones a la sustancia blanca cerca de la sustancia gris y constituyen los fascículos propios. Su interrupción produce trastornos en reflejos intersegmentarios. Son muy importantes en funciones reguladoras automáticas medulares como los que controlan la micción en segmentos lumbosacros o los que intervienen en el control sinérgico de los músculos respiratorios en la porción superior de la médula espinal.
La organización suprasegmentaria comprende los tractos ascendentes largos que llevan impulsos aferentes a centros encefálicos, y los tractos descendentes largos por los cuales estos centros superiores influyen en las motoneuronas inferiores.
Vías ascendentes o Sensitivas
Deben distinguirse las de la sensibilidad exteroceptiva o cutánea, aquellas de la sensibilidad propioceptiva o profunda y las de la sensibilidad interoceptiva o visceral.
Vías de la sensibilidad exteroceptiva o cutánea: se trata de una sensibilidad consciente, cuyo punto de partida se encuentra en los corpúsculos sensoriales de la piel, cada uno de los cuales posee una especialización precisa y según la especialización, tenemos las siguientes sensibilidades exteroceptivas: sensibilidad termoalgésica, táctil protopática y táctil epicrítica.
-
Sensibilidad termoalgésica: el axón de la primera neurona, que se encuentra en el ganglio raquídeo llega a la cabeza del cuerno posterior, o sea al área 1 del asta dorsal donde se pone en contacto con la segunda neurona, el axón de ésta atraviesa la línea mediana por la comisura gris, pasando detrás del cordón ependimario y llega al cordón lateral por donde va a ascender hasta el tálamo, para llegar hasta la tercera neurona que se encuentra en el núcleo ventoposterolateral, los axones de esta tercera neurona pasan a través del brazo posterior de la cápsula interna y la corona radiada, para alcanzar a la cuarta neurona en el área somatoestésica en la circunvolución poscentral de la corteza cerebral. La sensibilidad termoalgésica llega por el haz espinotalámico lateral.
-
Sensibilidad táctil protopática: haciende por el haz espinotalámico anterior, la primera neurona se encuentra en el ganglio raquídeo y su axón penetra en la sustancia gris, en el cuerno dorsal, específicamente en el área 1, tiene un trayecto muy similar al del haz espinotalámico lateral, y una diferencia es que el axón de su segunda neurona al pasar al lado opuesto, pasa por delante del conducto ependimario, luego asciende por el cordón anterior, hasta llegar a la tercera neurona que se encuentra en el tálamo, en núcleo ventroposterolateral y desde aquí los axones pasan a la corteza cerebral, a la circunvolución poscentral, a través de la cápsula interna y la corona radiada.
-
Sensibilidad táctil epicrítica: asciende hacia los centros superiores junto con los haces de la sensibilidad profunda consciente, siguiendo la siguiente vía, la primera neurona se encuentra en el ganglio raquídeo, su axón llega hasta los núcleos bulbares de Goll y de Burdach, asciende al tálamo con el lemnisco medial y ahí se encuentra la tercera neurona, cuyo axón a su vez sale del nucleo ventral posterior y llega hasta la corteza sensorial.
Vías de la sensibilidad propioceptiva o profunda: es la de las articulaciones y de los músculos, juega una acción muy importante en las actvidades musculares ayudando al sentido de actitud. Es una sensibilidad consciente e inconsciente. La sensibilidad será consciente siempre que llegue hasta la corteza sensitiva e inconsciente cuando no llegue hasta la corteza, a la sensibilidad inconsciente, la podemos clasificar en: sensaciones de presión o barestesia, de vibración o palestesia, y de actitud o posición que ocupa un miembro en el espacio o batiestesia.
-
Sensibilidad consciente: transmite sensaciones dolorosas. La primera neurona se encuentra en el ganglio espinal, su axón no pasa por el cuerno posterior; penetra directamente en el fascículo posterior del mismo lado, y asciende verticalmente hacia los centros superiores. En la cara dorsal de la médula un surco longitudinal, el colateral posterior, parece separar este fascículo posterior en dos porciones: la parte medial o fascículo de Goll, que lleva información sensitiva del tronco y del miembro inferior del mismo lado y la parte lateral, fascículo cuneiforme o de Burdach, que lleva información sensitiva del miembro superior y el cuello del mismo lado. La segunda neurona se encuentra en los núcleos de Goll y Burdach, emiten axones que se cruzan en la médula oblonga y que forman el lemnisco medial, al cruzar el puente, se unen a las fibras del haz espinotalámico anterior, la tercera neurona está en el núcleo ventroposterolateral del tálamo y la cuarta neurona, como es consciente en la corteza sensitiva, en la circunvolución poscentral, en el área somatoestésica.
-
Sensibilidad inconsciente: transmite esencialmente a los centros superiores, especialmente cerebelosos. La primera neurona se encuentra en el ganglio espinal, su axón termina en la parte media del cuerno dorsal, se articula allí con la segunda neurona situada en el núcleo torácico o columna de Clarke, para las neuronas que corresponden a los miembros inferiores y al tronco. El axón atraviesa enseguida la sustancia gris para formar en la parte posterior del fascículo lateral del mismo lado, sin cruzar la línea media, el tracto espinocerebeloso dorsal o fascículo espinocerebeloso de Flesching, las fibras provenientes de los miembros superiores, hacen contacto con el núcleo vestibular superior. El axón cruza entonces la línea mediana situada en la sustancia gris, alrededor del canal central y llega a la parte anterior del cordón lateral opuesto participando en la constitución del tracto espinocerebeloso cruzado o fascículo de Gowers.
Vías de la sensibilidad interoceptiva: transmiten la sensibilidad visceral. Sus vías son mal conocidas; al parecer siguen la sustancia gris de la médula.
Conclusión
La médula espinal tiene en escencia la misma disposición en toda su extensión. La sustancia gris, que presenta la forma de una letra H, se modifica localmente en función de las diferencias en el número y tipo de las neuronas que contiene. Estas son más abundantes en los abultamientos cervical y lumbar, ya que estas regiones inervan a los miembros. De manera similar, la sustancia blanca varía, aunque poco, en su disposición en la médula, y se encuentra contenida en tres cordones a cada lado. Estos cordones están formados por un conjunto de haces nerviosos que ascienden o descienden, según sean sensitivos o motores.
La vía motora, lleva información desde los centros superiores hasta donde se realizan los movimientos, los músculos, esta vía posee dos neuronas en su trayecto, podemos clasificar a las vías motoras en voluntarias o piramidales e involuntarias o extrapiramidales
La vía sensitiva, trae información que recoge de los receptores ubicados en la periferia e interior del cuerpo, esta información es transmitida por cuatro neuronas intermedias a lo largo de su recorrido y puede ser una información consciente, como el dolor, calor y tacto e inconsciente, si se trata de información proveniente de músculos y de tendones.
4. VIAS DESCENDENTES MEDULARES
Tractos descendentes largos
Existen diferentes niveles encefálicos que son capaces de influir en la actividad de las neuronas motoras inferiores de la médula espinal. El sistema piramidal (tractos corticoespinales) y extrapiramidal (tractos rubroespinal, vestibulo-espinal, reticuloespinal, tectoespinal) forman parte de estos niveles.
Tractos corticoespinales o piramidales
Son los tractos motores más importantes de la médula espinal, ya que conducen impulsos que inician y controlan los movimientos voluntarios. Debido a que esta vía eferente agrega rapidez y agilidad a los movimientos voluntarios, principalmente actúa en la realización de movimientos rápidos y hábiles. Las neuronas que constituyen los tractos corticoespinales se denominan neuronas motoras superiores. Sus fibras se originan en las células piramidales de la capa V de la corteza cerebral motora, aunque no exclusivamente. Las proporciones son las siguientes: un tercio de las fibras se origina en la corteza motora primaria, un tercio en la corteza motora secundaria (ambas regiones pertenecientes al giro precentral) y un tercio en el giro postcentral. Este último origen no controla la actividad motora precisamente, sino que envía impulsos sensitivos que modulan este tipo de respuestas.
Desde la corteza, alrededor de un millón de fibras pasa a formar parte de la corona radiada para luego penetrar en el brazo posterior de la cápsula interna. En este sitio, las fibras correspondientes a las regiones faciales están cercanas a la rodilla (genu) y las extremidades inferiores se sitúan en un aspecto más posterior de la cápsula interna. Las fibras continúan su camino constituyendo las 3/5 partes de la porción media de cada pie del pedúnculo cerebral. Las fibras correspondientes a la región cervical se ubican medialmente y las fibras de las extremidades inferiores lateralmente. En el puente, las fibras pontocerebelosas transversas separan el tracto en varios haces bien definidos. Al penetrar al bulbo raquídeo, las fibras vuelen a reagruparse para constituir las pirámides en la porción anterior de esta región. En la unión bulbomedular, las fibras se decusan parcialmente (decusación de las pirámides): La mayoría de las fibras (90%) pasan al lado opuesto y descienden en el cordón lateral como el tracto corticoespinal lateral, mientras una pequeña proporción no se decusa y desciende en el cordón anterior del mismo lado como el tracto corticoespinal anterior o directo.( Fig.6) A medida que estos tractos descienden por la médula, se hacen progresivamente más delgados debido a que sus fibras van haciendo conexiones con las motoneuronas de asta anterior en sucesivos niveles ya sea directa o indirectamente a través de interneuronas. Si bien las fibras del tracto piramidal directo no se decusan a nivel bulbar, sí lo hacen a través de la comisura blanca anterior en los segmentos cervicales y torácicos superiores para así alcanzar el asta anterior del lado opuesto. Las fibras del tracto corticoespinal lateral, ya decusadas a nivel bulbar, terminan en las astas anteriores de todos los segmentos medulares. A fin de cuentas, cada hemisferio cerebral se relaciona funcionalmente con el lado opuesto del cuerpo. Estudios histológicos experimentales han demostrado la presencia de fibras piramidales que conectan cada hemisferio cerebral con las motoneuronas inferiores ipsilaterales, pero parecen ser tan escasas que no son significativas frente a una valoración clínica.
La mayoría de las fibras corticoespinales sinapta con una interneurona, y es ésta la que se comunica con varias motoneuronas gamma, incluso con la de segmentos medulares adyacentes amplificando así la respuesta; sólo las fibras corticoespinales más largas son las que sinaptan directamente con las motoneuronas del asta anterior.
Existen colaterales de las fibras corticoespinales que sinaptan con los núcleos caudado, lentiforme, rojos, olivares y reticulares, con la función de mantener cierto flujo de información acerca de la actividad de la corteza motora. Estas regiones subcorticales podrían ejercer otras funciones reguladoras sobre las neuronas motoras inferiores a través de otras vías descendentes. Se sabe también de ramas corticoenucleares que se desprenden a nivel del tronco para contactar con los núcleos de los pares craneales.
Sistema extrapiramidal
Se ocupa el término de sistema extrapiramidal para referirse a los tractos descendentes que no sean los corticoespinales.
Tracto Rubroespinal
Sus fibras se originan en el núcleo rojo del mesencéfalo y transmiten impulsos que provienen de: (1) el mismo núcleo rojo e indirectamente de (2) cerebelo y (3) cuerpo estriado. Las fibras se decusan a la misma altura del núcleo rojo y continúan por el tronco encefálico hasta el cordón lateral de la médula espinal, anterior al tracto piramidal cruzado, para finalmente hacer sinapsis con las interneuronas de las astas anteriores. Se cree que es una importante vía indirecta por la cual el cerebelo y la corteza cerebral pueden influir sobre las motoneuronas inferiores alfa y gamma, facilitando la actividad de los músculos flexores e inhibiendo la actividad de los extensores. A medida que se asciende en la escala evolutiva, se observa una relación inversamente proporcional entre el desarrollo del tracto rubroespinal y los tractos piramidales: conforme los tractos piramidales se agrandan, el rubroespinal se adelgaza considerablemente. Esta es otra muestra más de la proscencefalización de las funciones, en donde los niveles funcionales superiores priman ante los inferiores. Todo lo antedicho explica por qué el tracto rubroespinal es pequeño y relativamente insignificante en el hombre.
Tracto Vestibuloespinal
Este tracto se presenta difusamente y mezclado con otros tractos en la periferia del cordón anterior de la médula espinal hasta sinaptar con interneuronas del asta anterior. Sus fibras, en mayoría homolaterales, se originan en el núcleo vestibular lateral del bulbo raquídeo, el cual recibe aferencias que provienen del sistema de canales semicirculares a través del nervio vestibular y del cerebelo. Las fibras terminan sinaptando con interneuronas de las astas anteriores. A través de este tracto, el oído interno y el cerebelo se encargan de facilitar la actividad de los músculos extensores e inhibir a los flexores, relacionándose así con la mantención del equilibrio.
Tracto Tectoespinal
Este tracto es mero remanente de una vía que controlaba las motoneuronas inferiores desde los centros de reflejos visuales mesencefálicos. Sus fibras se originan en el colículo superior del mesencéfalo y se decusan inmediatamente para descender por el tronco encefálico en una región cercana al fascículo longitudinal medial. En la médula espinal este tracto desciende a través del cordón anterior por delante del tracto reticuloespinal anterior cercano a la fisura mediana anterior. Sus fibras terminan sinaptando con interneuronas de los segmentos cervicales superiores. Está relacionado funcionalmente con los movimientos posturales reflejos producidos en respuesta a estímulos visuales.
Tractos Reticuloespinales
Estos tractos son importantes vías que llevan impulsos desde el mesencéfalo y rombencéfalo para controlar el tono muscular. Se originan en los difusos grupos de núcleos reticulares de la formación reticular del tronco encefálico. Los axones de los núcleos reticulares del puente descienden por el funículo anterior homolateral de la médula espinal formando el tracto reticuloespinal pontino. Los axones de los núcleos reticulares del bulbo descienden por el funículo lateral formando el tracto reticuloespinal bulbar. Gracias a su capacidad de inhibir o excitar las neuronas motoras inferiores alfa y gama de la columna gris anterior, es posible que este tracto participe en las siguientes funciones (1) control del tono muscular (2) ajuste de la respuesta cardiovascular por medio de neuronas del SNA (2) regulación de los movimientos respiratorios. Debido a su ubicación topográficamente mal definida, ha sido imposible obtener resultados experimentales que comprueben muchas de sus funciones.
Tracto Olivoespinal
Las fibras de este tracto se originan en el núcleo olivar inferior y descienden por el funículo lateral de la médula espinal para controlar la actividad de la motoneurona inferior.
5. PRINCIPIOS DE FISIOLOGÍA MEDULAR
Elementos de la placa motora:
Reflejo miotático:
Consiste en la contracción de las fibras extrafusales de un músculo, como resultado del estiramiento de las fibras intrafusales del mismo músculo. El reflejo se circunscribe al segmento medular donde se localizan las neuronas motoras del músculo estimulado y establece un arco monosináptico entre las gruesas fibras, las anuloespirales del huso y las neuronas alfa del asta anterior medular.
Reflejo de flexión:
Es el otro tipo de reflejo medular. Se desencadena por estímulos exteroceptivos, sobre todo la naturaleza nociva (nociceptivos), como pinchazos o quemaduras sobre la piel. La respuesta consiste en la flexión o retirada de los segmentos afectados. La respuesta de flexión no esta limitada a un solo segmento medular, sino que abarca varios segmentos por encima y por abajo del segmento dermatomico estimulado, debido a la distribución por varios segmentos de una solo fibra exteroceptiva, por lo cual el reflejo también ha recibido el nombre de reflejo multisináptico.
Reflejo de extensión invertido:
El órgano músculo - tendíneo de Golgi es un defensor del músculo a la tensión exagerada de sus fibras. Sus terminaciones tienen un alto dintel de excitación, mayor que en el de los husos, por lo cual solo descargan sus estímulos con tensiones musculares mucho mayores que las requeridas para desencadenar el reflejo miotático, pero una vez alcanzado esta nivel critico, sus estímulos anulan la contracción muscular por inhibición de la neurona alfa a través de sinapsis con células inhibitorias. Esta acción se ha conocido como reflejo de extensión invertido.
Tono muscular:
Es la contracción sostenida e involuntaria del músculo, debida a la acción nerviosa y que no origina movimientos o cambios de posición de las articulaciones.
Las contracciones musculares no siempre producen un movimiento real. En la mayoría de los músculos existe una pequeña fracción del numero total de fibras que se contraen en forma continua. Esto permite mantener la postura de una extremidad y la capacita para resistir la elongación o el estiramiento pasivo. Esta leve contracción mantenida se denomina tono muscular.
Características de la neurona motora superior e inferior:
Se conoce que la neurona inferior se le denomina gamma, debido a su bajo umbral de estimulación, y la superior se le llama alfa. De acuerdo a estas dos se adecua el tono del músculo con la finalidad del movimiento que se va ejecutar.
La acción de la neurona gamma, asegura la permanencia de la actividad refleja del tono durante y después de la contracción muscular.
6. ANATOMIA EXTERNA DEL TALLO CEREBRAL
6. ANATOMIA EXTERNA DEL TALLO CEREBRAL
Porciones del tallo cerebral:
Esta configurado por los derivados del diencéfalo, mesencéfalo, metencéfalo y mielencéfalo embrionario; de los cuales tres son de gran importancia:
Bulbo raquídeo.
Protuberancia o puente.
Mesencéfalo.
Superficie ventral:
Esta formada por un grueso cuerpo de fascículos nerviosos transversalmente dispuestos, el Brachium pontis o pedúnculo cerebeloso medio, que dirigido dorsalmente y cubriendo el cuerpo restriforme va a terminar al cerebelo. En la parte ventral, el pedúnculo cerebeloso medio presenta un surco longitudinal, amplio y poco profundo, en el cual se aloja la arteria basilar.
Superficie dorsal:
La superficie posterior del bulbo enmarcada por los cuerpos restriformes se denomina como el piso del cuarto ventrículo.
Cuarto ventrículo:
La dilatación de la tercera vesícula cerebral primaria del embrión (rombencéfalo) evoluciona como el cuarto ventrículo. Esta cuarto ventrículo es una cavidad situada entre el bulbo y la protuberancia ventralmente y el cerebelo en su porción dorsal. Caudalmente se continua con el canal central del bulbo y la medula, y rostralmente con un delgado conducto, el acueducto de Silvio, en el espesor del mesencéfalo.
7. ANATOMIA INTERNA DEL BULBO RAQUIDEO
Pirámides bulbares: Son gruesos fascículos en la cara vertical del bulbo, forrados por fibras corticoespinales y corticobulbares, las fibras corticoespinales pasa al cordón lateral del lado opuesto en la decusación de las pirámides, y forma así el fascículo piramidal cruzado
Lemnisco medio: Banda ascendente de fibras formada por los axones de las células de los fascículos graciulis y cuneatus; esta detrás de las pirámides y termina haciendo contacto con las neuronas del nuc
leo ventral posterolateral del tálamo
Núcleos olivares: El núcleo olivar inferior es una gran lamina de substancia gris irregularmente plegada sobre si misma, de tal manera que su concavidad o hilio mira hacia la línea media. En el se encuentran los núcleos olivares accesorio dorsal y medial. Los diversos núcleos olivares están formados por neuronas de mediano tamaño previstas de dendritas cortas y un delgado axon.
Núcleos arciformes: Son pequeñas masas nucleares situadas en la porción dorsomedial de las pirámides.
Núcleo motor dorsal del vago: Se extiende por fuera del núcleo del hipogloso, desde el piso del cuarto ventrículo, al ala cineárea. De las células de este núcleo se originan en las fibras que enervan la musculatura lisa y el tejido glandular.
8. ANATOMIA INTERNA DE LA PROTUBERANCIA
Porción basilar de la protuberancia: Esta formada por fibras y verticales y transversales. En medio de estos haces de fibras se encuentran los núcleos de la corteza cerebral, en ellas se pueden diferenciar varios grupos: fibras corticoespinales, fibras corticoespibares y fibras corticopónticas. Las fibras horizontales forman los fascículos constitutivos del brachium pontis, que a su vez constituyen el pedúnculo cerebeloso medio.
Porción tegmental: Se presenta en ella algunas de las estructuras apreciables en el bulbo, como la formación reticular y las vías ascendentes y entre estas el lemnisco medio.
Composición funcional
Núcleos principales
Núcleo del motor ocular externo: Se localiza al nivel de coliculo facial, engrosamiento de la eminencia media producido por el núcleo del motor ocular externo y la rodilla del nervio facial.
Núcleo motor del facial: Esta situado en la porción tegmental y en la mitad inferior de la protuberancia. Esta formado por grupos de neuronas motoras cuyos axones forman un fascículo compacto que se dirige del cuarto ventrículo.
Núcleos del trigémino: Son los núcleos motor y sensitivo del V par, se encuentran en la porción tegmental, y al la altura del tercio superior de la protuberancia.
Núcleos cocleares (relacion y conexiones): Los núcleos cocleares, dorsal y ventral, forman el coliculo acústico, que rodea el cuerpo restiforme. Las fibras de los núcleos cocleares se dirigen hacia la zona limite entre la porción tegmental y la porción basilar donde forman el cuerpo trapezoide. Los núcleos cocleares reciben las impresiones sonoras que vienen de los nervios cocleares. Los axones de las celulas de los núcleos forman el lemnisco lateral.
9. ANATOMIA INTERNA DEL MESENCEFALO
Porción dorsal y basal: El mesencéfalo esta constituido por dos porciónes una basal, pedúnculo cerebral y una dorsal tectum o lamina cuadrigemina, en la cual se encuentran los coliculos cuadrigeminos anteriores y posteriores. Pedunculo cerebral: tiene dos porciónes , una ventral o base o pie del pedunculo y una porción dorsal o tegmentum; en la base del pendunculo se encuentra la fosa interpendular y en el tegmentum se encuentra la substancia nigra, que es un gran núcleo.
Acueducto de silvio: Se encuentra en la lamina cuadrigemina, es un delgado conducto que comunica el tercero con el cuarto ventrículo y esta rodeado por una gruesa capa de substancia gris.
Núcleos principales
Substancia nigra: Es una lamina gruesa de substancia gris que se extiende desde el borde inferior del mesencefalo hasta el diencefalo.
Núcleo rojo: Esta en la parte dorsal medial de la substancia nigra, es in masa redondeada de substancia gri, se extiende en longitud desde el borde ingerior del coliculo cudrigemino inferior hasta el subtalamo. Tiene conexiones con diversos núcleos del tallo cerebral y el núcleo olivar.
Núcleo del patetico: es una pequeña masa situada a la altura del coliculo cuadrigemino inferior a nivel del borde anterior de la substancia gris central. Este núcleo da inervacio al músculo oblicuo superior del ojo.
Núcleo del motor ocular comun: Se encuentra a nivel del borde anterior o ventral de la substancia gris central. Ls celulas del núcleo motor ocular comun pueden separarse en grupos del recto externo y del oblicuo superior.
Núcleo de Edinger-Westphal: Esta situado en la parte rostral y sobre la línea media, de aquí los impiulsos pasan al ojo a traves de los nervio ciliares que dan la inervacion al musculo ciliar y al musculo constricto de la pupila.
Núcleo mesencéfalico del trigémino: formado por un grupo de celulas ovoides, de tipo sencitivoque rodean lateralmente la sustancia gris central del mesencefalo. Por medio de él se llevan las sensaciones musculares y articulares de la cabeza hasta el tálamo, a través del lemnisco trigéminal.
Síndromes por lesiones del bulbo raquídeo, protuberancia y mesencéfalo
Síndromes bulbares:
- Síndrome ventral del Bulbo
- Síndrome de Wallemberg
- Síndrome de Avellis
- Síndrome de Schmidt
- Síndrome de Jackson
- Síndrome de Tapia
- Síndrome de Cestan Chenais
Síndromes protuberanciales:
- Síndrome del ángulo pontocerebeloso
- Síndrome de Millard-Gubler
- Síndrome de protuberancia inferior
- Síndrome de Raymond Cestan
Síndromes de Mesencéfalo:
- Síndrome de Weber
10- CLASIFICACION FUNCIONAL DE LOS NUCLEOS RAQUIDEOS Y CRANEAES
El sistema nervioso se divide en dos grandes partes:
*SISTEMA NERVIOSO SOMATICO: por medio del cual tenemos las sensaciones del ambiente que nos rodean y reaccionamos ante el por medio de nuestro sistema muscular esquelético.
*SISTEMA NERVIOSO VEGETATIVO: relacionado con el funcionamiento visceral.
Estas divisiones funcionales dan cuatro tipos de estructura:
Somáticas motoras o somáticas eferentes: dan inervaciòn a la musculatura estriada esquelética.
Somáticas sensitivas o somáticas aferentes generales: perciben las sensaciones de los receptores cutáneos ( exterocepcion) y de los músculos y articulaciones (propiocepcion)
Viscerales motoras o viscerales eferentes generales: encargadas de la inervacion motora de las vísceras
Visceral aferente general: percibe la sensibilidad visceral.
Se les da el calificativo de generales por estar distribuidas en todo el organismo.
NUCLEOS PRINCIPALES
1-GRUPO SOMATICO EFERENTE
Núcleo del motor ocular común.
Núcleo del patético
Núcleo del motor ocular externo
Núcleo del hipogloso.
Estos núcleos dan inervaciòn a músculos derivados de somites.
2-GRUPO VISCERAL EFERENTE GENERAL
Núcleo dorsal del vago
Núcleo lacrimal
Núcleo salivatorio superior e inferior
Núcleo de Edinger-Westphal
Las fibras eferentes de estos núcleos son preganglionares y hacen sinapsis con neuronas de los ganglios periféricos del sistema nervioso vegetativo y dan inervacion al músculo liso, al músculo cardiaco y al tejido glandular.
3- GRUPO VISCERAL EFERENTE ESPECIAL
Núcleo ambiguo
Núcleo motor del facial
Núcleo motor del trigémino
El ambiguo encargado de la inervaciòn a los músculos de la laringe y la faringe, el núcleo motor del facial que inerva los músculos faciales y el núcleo motor del trigémino que da la inervaciòn a los músculos masticadores.
4- GRUPO SOMATICO AFERENTE GENERAL
Sensibilidad exteroceptiva: Núcleo de la raíz descendente y núcleo sensitivo principal del trigémino.
Sensibilidad propioceptiva: Núcleo mesencefalico del trigémino.
Es el núcleo de origen de las fibras propioceptivas de los músculos masticadores y articulación temporomaxilar.
5- GRUPO SOMATICO AFERENTE ESPECIAL
Núcleos cocleares, dorsal y ventral
Núcleos vestibulares, descendentes, medial, lateral y superior.
Las fibras del nervio vestibular se originan de las células del ganglio vestibular y las del auditivo en las células del ganglio espiral de la coclea.
6- GRUPO VISCERAL AFERENTE GENERAL
Núcleo del fascículo solitario: conformado por neuronas del ganglio geniculado y del ganglio petroso, para dar sensibilidad al oído medio, vasos de la cara y vasos intracraneales; la base de la lengua, la faringe, la trompa de eustaquio, el oído medio y el seno carotideo.
7- GRUPO VISCERAL AFERENTE ESPECIAL
Núcleo del fascículo solitario (porción rostral)
Este grupo corresponde a las vías nerviosas conectadas con el sentido del gusto.
MANIFESTACIONES CLINICAS DE LESIONES DEL TALLO CEREBRAL
Las manifestaciones clínicas de las lesiones del tallo cerebral son comprensibles y predecibles si se tiene en cuenta su organización estructural.
Algunos signos dependen de lesiones de las vías sensitivas o motoras y otras de las lesiones de los núcleos o fibras de los nervios craneales.
SINDROMES BULBARES: se esquematizan en tres grupos:
-
Producidos por lesiones de la porción ventral
-
Producidos por lesiones de la región lateral del bulbo
-
Producidos por lesiones de la región dorsal
SINDROME VENTRAL DEL BULBO (Síndrome de Reyniold-Revillod y Dejerine)
Hay compromiso de una pirámide y de las raíces del hipogloso lo que produce una hemiplejía contralateral y una parálisis con atrofia de la mitad de la lengua del mismo lado.
SINDROME DE WALLENBERG: Es el mas frecuente de los síndromes del bulbo. Se presenta parálisis del velo del paladar, de la faringe y de la cuerda vocal homolateral debido al compromiso del núcleo ambiguo. Hay perdida de la sensibilidad gustativa en la mitad de la lengua. El síndrome es debido generalmente a trombosis de la arteria cerebelosa posterior inferior.
SINDROME DE AVELLIS: producido por trombosis de la arteria vertebral. Se produce parálisis del velo del paladar y de la cuerda vocal del mismo lado, además, perdida de la sensibilidad al dolor y la temperatura en el lado opuesto del cuerpo.
SINDROME DE SCHMIT: además de las alteraciones del anterior hay parálisis del trapecio y del esternocleidomastoideo.
SINDROME DE JACKSON: a mas del cuadro del síndrome de Schmit, hay compromiso del hipogloso, lo que produce atrofia lingual.
SINDROME DE TAPIA: es producido por lesiones de la región dorsal de bulbo; hay parálisis ipsilateral de la faringe y la laringe y parálisis con atrofia de la lengua.
SINDROME DE CESTAN CHENAIS: es producido por trombosis de la arteria vertebral, con destrucción casi completa del bulbo. Similar su sintomatologia a la del síndrome de Babinski.
SINDROMES PROTUBERENCIALES:
Con compromiso de los nervios VII y VIII, y a lesiones del de la región basilar.
SINDROME DEL ANGULO PONTOCEREBELOSO: se debe principalmente a la compresión del VIII par. Hay perdida de la función auditiva de tipo nervioso con paralisis de tipo periferico.
SINDROME DE MILLARD-GUBLER: Hay parálisis homolateral de la cara y hemiplejía contralateral; puede haber también extravismo interno y parálisis del ojo homolatera.
SINDROME PROTUBERENCIAL SUPERIOR: produce parálisis del motor ocular externo homolateral. En algunos casos hay parálisis de los movimientos conjugados de los ojos.
SINDROME DE RAYMOND-CESTAN: se manifiesta como hemiplejía y anestesia cruzadas y una parálisis del recto externo homolateral, asociado a trastornos como, temblor, ataxia y asinergia del mismo lado de la lesión
SINDROMES RELACIONADOS CON EL MESENCEFALO:
Se cataloga según su localización en tres grupos:
-
Producidos por lesiones del pie del pedúnculo
-
Producidos por lesiones de la region tegmental
-
Producidos por lesiones del tectum
12)SINDROME DE WEBER: Se caracteriza por una hemiplejía contralateral y parálisis ipsilateral del III par, se manifiesta por ptosis palpebral, parálisis y desviación externa del lobo ocular y dilatación de la pupila.
13)SINDROME DE BENEDIKT: Como en el anterior hay parálisis, a lo cual se agrega sintomatologia secundaria a la lesión del núcleo rojo, consiste en temblor, ataxia e hiperquinesia del lado opuesto del cuerpo.
14) SINDROME DE PARINAUD: se caracteriza por una perdida del movimiento conjugado de los ojos hacia arriba.
11.SISTEMA NERVIOSO AUTONOMO Y VEGETATIVO
Definir el SNA. Diferencias con el sistema somático:
En el Sistema Somático, las estructuras efectoras (fibras musculares estriadas ) reciben la inervación directamente de los centros nerviosos cerebroespinales.
En el sistema vegetativo la inervación de las estructuras efectoras (músculos involuntario, glándulas ) se establea través de una cadena de dos neuronas, una central, localizada en los órganos cerebroespinales y una periférica localizada en uno delos ganglios periféricos del sistema .
Definir sistemas nervioso simpático y parasimpático características anatómicas:
Sistema nervioso simpático son fibras preganglionares o presinápticas , se originan en la columna mediolateral de la médula espinal (astas laterales ).
Sistema nervioso parasimpático fibras preganglionares se originan en núcleos del tallo cerebral y en la región sacra de la médula espinal.
| ORGANO | PARASIMPATICOS | SIMPATICO |
| Pupila | Contracción | Dilatación |
| Corazón | Retardo | Aceleración |
| Bronquios | Contracción | Dilatación |
| Esfínteres Intestinales | Relajación | Contracción |
| Vasos Coronarios | No Definido | Dilatación |
| Vasos De Piel Y Mucosa | No Definido | Constricción |
| Vasos De Esqueletico | Dilatación | Constricción |
| Glándulas Sudoriparas | Secreción Generalizada | Secreción Local |
| Glándulas salivares | Secreción Acuosa | Secreción Viscosa |
| Vejiga | Contracción del Detrusor | Contracción incompleta del Detrusor |
| Tubo Digestivo | Aumento del peristaltismo | Disminución Del Peristaltismo |
| Arterios Helicineas | Dilatación |
Definir Núcleos Principales, fibras Aferentes y Eferentes:
FIBRAS EFERENTES
Núcleos de Edinger- Westphal:
Las fibreas de células de este ganglio salen con las del III para terminar en el ganglio ciliar. De este salen fibres postganglionales que pasan al globo ocular, haciendo parte del los nervios ciliares cortos y terminan en el músculo ciliar y en el músculo constrictor de la pupila.
Núcleo Salivatorio Superior :
Situado en la formación reticular pontobulbar, sus fibras salen con el intermediario de wrisberg, pasan luego a la cuerda del tímpano y al nervio lingual para terminar en el ganglio submaxilar.Estas fibras se distribuyen en las glándulas submaxilar y sublingual.
Núcleo Lacrimal :
Aproximadamente con la misma localización que el núcleo salivatorio superior se encuentra el núcleo lacrimal cuyos axones salen con las fibras del intermedio. Las fibras eferentes de este ganglio se unen a al división maxilar del trigémino ;siguen sus ramas zigomáticas orbitiaria de la que se desprenden para unirse a la rama lacrimal del oftálmico y llegar a al glándula lacrimal.
Núcleo salivatorio inferior:
También situado en la región pontobulbar . Sus fibras salen con las del glosofaríngeo, se desprenden de él nervio timpánico, atraviesan el plexo timpánico y salen del temporal como el nervio petroso superficial menor que termina en el ganglio ótico sus fibras postganglionares llegan a la parotida a través del nervio auriculo temporal, rama del v par.
Núcleo Motor dorsal del vago:
Situado en el piso de la fosa romboidal.Sus fibras dan la inervación parasimpática a las vísceras torácicas y a la mayoria de las vísceras abdominales.Son fibras preganglionares que terminan haciendo sinapsis con las células de los ganglios terminales existentes en contacto con las vísceras incluidas en el interior de ellas.Se distribuyen en:
Nervios Cardíacos: Situado en el epicardio de la aurícula izquierda y de los apndíces auriculares y con los nervios cardíacos del simpático, forman los plexos cardíacos las fibras postganglionares terminan en la musculatura auricular y en las fibras del seno y del nodo del sistema de
Conducción cardíaca.
Ramos Pulmonares:Presentes en la pared bronquial, cuyos axones, fibras postganglioneres, terminan en contacto con la musculatura lisa bronquial.
Ramos esofágicos, ramos gástricos:provenientes de ambos vagos.
Ramos intestinales: Se origina especialmente del vago derecho, llegan al intestino delgado y a la porción proximal del grueso, las fibras postganglionales de estos plexos regulan tanto la contractilidad de la musculatura lisa intestinal como la secreci´´on de sus glándulas.
Ramos para vesícula biliar y conductos víliares: fibras del neumogástrico que hacen parte del plexo celíaco, llegan a los órganos biliares a través del ligamento gastrohepatico. Inerva la musculatura lisa de estos conductos.
VIAS AFERENTES
Facial: se cree que este nervio conduce la sensibilidad profunda y vísceral general de la cara, oido medio, trompa de eustaqui, pared faríngea y glándulas salivares. Las axones forman la raíz sensitiva del facial, o nervio intermedio de wrisberg.
Glosofaríngeo :las fibras sensitivas viscerales generales de este nervio reciben los estímulos de la porción posterior de la lengua, las admídalas, la pared faringeas, y las trompas de eustaquio; ellas constituyen ramas faringeas, linguales y tímpanicas de las neuronas del ganglio petroso.
Neumogastrico: El X par craneal tiene numerosísimas fibras sensitivas que recogen distintos tipos de sensibilidad ;muchas de ellas se distribuyen a todo largo del tubo digestivo, de la raíz de la lengua hasta la parte media del colon transverso; otras al árbol respiratorio desde la laringe, hasta los alvéolos pulmonares y otras al arco de la aorta y la pared de la aurícula derecha.Estas fibras llevaran hasta los centros superiores de integración noticia del estado funcional de los aparatos digestivo, respiratorio y vascular para lograr la acomodación funcional a las exigencias del momento.
Fibras preganglionares : los axones de la células de la columna medio lateral de los segmentos S2, S3 y S4 salen de la médula y constituyen los llamados nervios pélvicos, se juntan con las ramas de los últimos nervios de la cadena símpatica (nervio presacro para formar el plexo hipogástrico que controla el funcionamiento de los órganos genitales, vejiga , uretra y la parte distal del colon.
Conexiones centrales reflejas de las vís viscerales aferentes:muchos de sus axones hacen contacto con el núcleo motor dorsal del vago lo que explica la existencia de reflejos vago-vagales : con los núcleos del nervio hipogloso; conel núcleo ambiguo para integrar así el reflejo de la deglución ; y especialmente , con grupos nucleares de la formació reticular.
12. CEREBELO
Determinar Areas anatómicas, anatomía interna:
Núcleo Dentado: es una lámina grandemente plegada y convocada hacia la línea media, situada en la parte anterior y medial del cuerpo medular;lateralmente se extiende bastante en el interior de los hemisferios.
Núcleo Boliforme:es una delgada lámina de substancia gris situada medialmente al dentado.
Núcleo Globoso: Está constituido por varias masas grises situadas por dentro del enboliforme. Por sus conecciones los anteriores núcleos estan en relacio´n con ,las estructuras del neo y paleocerebelo.
Núcleo Fastigiales: están situados en la parte anterior del cerebelo en la línea media, inmediatamente encima del techo del IV ventrículo y es el mas interno del núcleo cerebeloso; sus conecciones se hace principalmente con el archicerebelo y los núcleos vestibulares.
Anatomia Interna: la sustancia gris forma una capa continua o crteza que recubre las laminillas .La sustancia o cuerpo medular, forma el espesor del órganao y está formado por fibras que salen o entran a través de los pedúnculos. Por su aspecto ramificado, el cuerpo medular en loscortes sagitales del cerebelo, ha recibido el nombre del árbol de la vida (árbol vital).
Conexiones Aferentes y Eferentes:
Aferentes. Fascículo espinocerebeloso dorsal: su fibra se origian en el núcleo de clark de la medula espinal y terminan como fibras musgosas en el lóbulo anterior en la uvula y en la pirámide.
Fibras olivocerebelosas: Los distintos núcleos de complejo olivario exterior tienen un distinto significado filogenético que se reflejan en sus conexiones con el cerebelo. La mayor evidencia señala que las fibras olivocerebelosas terminan como fibras trepadoras en la corteza del cerebelo.
Fibras Reticulocerebelosas: se originan en varios núcleos reticulares del bulbo y del puente, entre ellos el núcleo reticular lateral y el paramediano del bulbo y el núcleo tegmental del puente.
Fibras arciformes externas dorsales o fascículo neocerebeloso. Se originan en los núcleos cuneatos.Conducen estímulos propioceptivos y táctiles del cuello y del miembro superior y tienen en el cerebelo una distribución similar a los fascículos cerebelosos.
Fibras arciformes externa ventrales y estías medulares del piso del cuarto ventrículo .vienen los núcleos arcutos del bulbo que se cnsideran una continuación de los núcleos pónticos.
Fascículos vestibulocerebelosos : son unos directos, originados en el ganglio de scarpa y otros indirectos que parten de los núcleos vestibulares.Terminan en la corteza del lóbulo floculonodular, en la ´vula y en núcleo fastigial.
Fibras trigeminocerebelosas: se origina en los cielos sensitivos del trigémino y llevan al cerebelo estimulos provinientes de la cabeza.
Eferentes. Forman mayor volumen del pedúnculo. En la unión del mes encéfalo con la protuberancia las fibras penetran al tegmento mesencefalico para cruzarse y terminar unas en el núcleo rojo y otras, de curso ascendente, en el núcleo ventral lateral del tálamo .
Importancia De la Coordinación Motora,tono moscular y equilibrio.
Control del equilibrio. Se observa principalmente como consecuencia de un tumor, en meduloblastoma, en la parte media del techo del cuarto ventrículo y que compromete las porciones del archi y el paleocerebelo. Y marcada inseguridad para el mantenimiento de la posición erguida, con tendencia con caer hacia tras
Equilibrio.Archicerebelo: pérdida de equilibrio, aumenta base sustentación, vértigo, reflejos anómalos.
Tono muscular: bajo o alto (hipotónico o hipertónico).
Coordinacion Motora. Neocerebelo: Ataxia (en coordinación motora) no se mide distancia, fuerza, dirección de movimientos
13. HEMISFERIOS CEREBRALES
Definir fisuras, surcos, circunvoluciones y lóbulos cerebrales.
Fisuras: Interemisferica separan los dos hemisferios cerebral. In situ, la fisura esta ocupado por un repligue de la dura madre, la hoz del cerebro. Si separan los bordes de la fisura Interhemisférica se descubre una gruesa banda de substancia blanca que une ambos hemisferios y que recibe el nombre de cuerpo calloso.
Surcos Precentral: está situado un poco por delante de la fisurade Rolando y paralelo a ella; entre los dos se delimita el giro precentral.
Surco frontal superior: se extiende por la superficie medial del hemisferio hasta la fisura callosomarginal.
Surco frontal inferior: se encuentra dividido en tres porciones por las ramas horizontal anterior y ascendente anterior de la fisura lateral.
Circunvolución: parietal inferior de distinguen dos porciones, el giro supramarginal, qu bordea el extremo posterior de la fisura lateral y el giro angular, que forma el reborde de la extremidad posterior del surco temporal superior.
Lóbulos Cerebrales: con base en las anteriores estructurales podemos dividir la superficie cerebral en varios lóbulos:
Lóbulo frontal: situado por delante del surco central y por encima de la fisura lateral y su rama horizontal posterior.
Lóbulo Parietal: Situado por detrás del surco central, se extiende hasta una línea que en la superficie externa corresponde a la proyección de la fisura parieoccipital, por debajo está limitado por la prolongación horizontal posterior de la fisura lateral.
Lóbulo Occipital: comprende el polo posterior del hemisferio, parietooccipita.
Lóbulo Temporal: en la superficie lateral del hemisferio situado por debajo dela fisura lateral y su prolongación horizontal posterior;en la superficiemedial avanza hasta contactarel diencéfalo.
Localizaciones Anatómicas
Un estudio que combina los registros neurofisiológicos (microelectrodos) con la histología de la corteza cerebral, sugiere que la corteza esta organizada en unidades verticales de actividad funcional.
Área Frontal
Área Motora Primaria: se extiende sobre le limite superior del lobulillo paracentral. Si se estimula produce movimientos aislados en el lado opuesto del cuerpo y contracción de grupos musculares relacionados con la ejecución de un movimiento específico. Las áreas del cuerpo están representadas en forma invertida en la circonvolución precentral. Comenzando desde abajo hacia arriba: deglución, lengua, maxilares, labios, laringe, párpado y cejas, dedos, manos, muñeca, codo, hombro y tronco etc.
La función del área motora primaria consiste en llevar a cabo los movimientos individuales de diferentes partes del cuerpo. Como ayuda para esta función recibe numerosas fibras aferentes desde el área premotora, la corteza sensitiva, el tálamo, el cerebelo y los ganglios básales. La corteza motora primaria no es responsable del diseño del patrón de movimiento sino la estación final para la conversión del diseño en la ejecución del movimiento.
Área Pre-motora: no tiene células gigantes de Betz. La estimulación eléctrica de esta zona produce movimientos similares a los del área motora primaria pero se necesita estimulación más intensa para producir el mismo grado de movimiento. Recibe numerosas aferencias de la corteza sensitiva, tálamo y ganglios básales. La función de ésta área es almacenar programas de actividad motora reunidos como resultado de la experiencia pasada; es decir programa la actividad motora primaria.
Área Motora Suplementaria: se ubica en la circunvolución frontal medial y por delante del lobulillo paracentral. La estimulación de esta área dá como resultado movimientos de las extremidades contralaterales pero es necesario un estímulo más fuerte que el necesario en la zona primaria. La eliminación de ésta área no produce una pérdida permanente de movimiento.
Campo Ocular Frontal: se extiende hacia delante desde el área facial de la circunvolución precentral hasta la circunvolución frontal media. La estimulación de esta área produce movimientos conjuntos de los ojos en especial en el lado opuesto. Controla los movimientos de seguimiento voluntario de los ojos y es independiente de los estímulos visuales. El seguimiento involuntario ocular de los objetos en movimiento comprende el área visual en la corteza occipital que está conectada al campo visual en la corteza occipital que está conectada al campo ocular frontal por fibras de asociación.
Área Motora del Lenguaje de Broca: está ubicada en la circunvolución frontal inferior entre las ramas anterior y ascendente y las ramas ascendente y posterior de la cisura lateral. En la mayoría de los individuos esta área es importante en el hemisferio izquierdo o dominante y su ablación da como resultado parálisis del lenguaje. La ablación de la región en el hemisferio no dominante no tiene efectos sobre el lenguaje. Produce la formación de palabras por sus conexiones con las áreas motoras adyacentes, músculos de la laringe, boca, lengua etc.
Corteza Pre-frontal: ocupa la mayor parte de las circunvoluciones frontal superior, media e inferior. Está vinculada con la constitución de la personalidad del individuo. Regula la profundidad de los sentimientos y está relacionada con la determinación de la iniciativa y el juicio del individuo.
Área Parietal
Área Somatoestésica Primaria: ocupa la circunvolución postcentral sobre la superficie lateral del hemisferio y la parte posterior del lobilillo paracentral sobre la superficie medial. Histológicamente es un área de tipo granuloso con capa externa de Ballinger muy ancha y obvia. La mitad opuesta del cuerpo está representada de forma invertida: faringe, lengua, cara,..., dedos, mano, brazo, tronco, muslo,.., pierna , pie. La porción de una parte del cuerpo en particular se relaciona con su importancia funcional y no con su tamaño. Por ejemplo superficies grandes ocupan la mano, la cara, labios y el pulgar.
Aunque la mayoría de las sensaciones llegan a la corteza desde el lado contralateral del cuerpo, algunas provenientes de la región oral van en el mismo sentido.
Área Somatoestésica de Asociación: ocupa el lobulillo parietal superior que se extiende hacia la superficie medial del hemisferio. Tiene muchas conexiones con otras áreas sensitivas de la corteza. Se cree que su principal función consiste en recibir e integrar diferentes modalidades sensitivas. Por ejemplo reconocer objetos colocados en las manos sin ayuda de la vista, es decir maneja información de forma y tamaño relacionándola con experiencias pasadas.
Área Occipital
Área Visual Primaria: ubicada en las paredes de la parte posterior del surco calcarino ocasionalmente alrededor del polo occipital. Histológicamente es un área de corteza delgada, del tipo granuloso con sólo algunas células piramidales. Recibe fibras que vienen de la retina. La mácula lútea, área central de la retina (área de la visión más perfecta) está representada en la corteza en la parte posterior. Las partes periféricas de la retina están representadas por el área anterior.
Área Visual Secundaria: rodea el área visual primaria. Recibe fibra aferentes del área visual primaria y otras áreas corticales y el tálamo. La función consiste en relacionar la información visual recibida por el área visual primaria con experiencias visuales pasadas, lo que permite reconocer y apreciar lo que se está viendo.
Se cree que existe un campo ocular occipital en el área visual secundaria cuya estimulación produce la desviación conjugada de los ojos cuando está siguiendo a un objeto, movimientos involuntarios que dependen de los estímulos visuales.
Área Temporal
Área Auditiva Primaria: está ubicada en la pared inferior del surco lateral. Histológicamente de tipo granuloso, es un área de asociación auditiva. La parte anterior del área auditiva primaria está vinculada con la recepción de sonidos de baja frecuencia mientras que la parte posterior con los de alta frecuencia. Una lesión unilateral produce sordera parcial en ambos oídos con mayor pérdida del lado contralateral.
Área Auditiva Secundaria: ubicada detrás del área auditiva primaria. Se cree que esta área es necesaria para la interpretación de los sonidos.
Área Sensitiva del Lenguaje de Wernicke: está ubicada en el hemisferio dominante izquierdo, principalmente, principalmente en la circunvolución temporal superior. Está conectado con el área de Broca por el haz de fibras llamado fascículo arcuato. Recibe fibras de la corteza visual (occipital) y de la corteza auditiva (temporal superior). Permite la compresión del lenguaje hablado y de la escritura, es decir que uno pueda leer una frase, comprenderla y leerla en voz alta.
Otras:
Área del gusto: está ubicada en el extremo inferior de la circunvolución postcentral de la pared superior del surco lateral en el área adyacente de la ínsula.
Área vestibular: está situada cerca de la parte de la circunvolución postcentral vinculada con las sensaciones de la cara. Su localización opuesta al área auditiva.
Insula
está enterrada dentro del surco lateral y forma su piso. Histológicamente es granulosa. Sus conexiones se conocen en forma incompleta se cree que se asocian con las funciones viscerales.
Todas las áreas restantes, tienen seis capas celulares y se conocen como áreas de asociación. Antes se suponía que recibían información provenientes de áreas sensitivas primarias, la integraban ya la analizaban, esto no ha sido comprobado. Actualmente se cree que tienen relación con la conducta, la discriminación y la interpretación de experiencias sensitivas.
El área asociativa frontal desempeña un papel que tiene que ver con las experiencias sensitivas pasadas, por ejemplo recordar una música escuchada en el pasado.
En el área asociativa parietal posterior, se integran aferencias sensitivas de tacto y presión, es decir tamaño, forma, textura. Esta capacidad se conoce como esterognosia. También se forma la apreciación de la imagen corporal.
Dominancia cerebral: si bien las circunvoluciones y las cisuras corticales son casi idénticas y es más, las vias que se proyectan también, ciertas actividades nerviosas son realizadas predominantemente por uno de los dos hemisferios cerebrales. La destraza manual, la percepción del lenguaje y el habla están controlados por el hemisferio dominante (en 90% de la población el izquierdo). Por el contrario la percepción espacial, el reconocimiento de las caras y la música por el no-dominate.
Se cree que en el neonato los dos hemisferios tienen capacidades equipotenciales. Durante la primer infancia en hemisferio domina al otro y sólo después de la primera década de vida la dominancia queda establecida.
Surcos y Giros De La Ínsula. Delimitan al surco circular y radian unas series de surcos distribuido en dos grupos anterior y posterior, separado por uno mayor el surco central de la insufla; son cortos y delimitan los cortos breves o cortos de la ínsula; los posteriores en numero de dos o tres, son mas largos y delimitan los giro largos de la ínsula. (Giro del cíngulo, hipocampo, fusiforme y ligual).
14. LINEA MEDIA CEREBRAL
Cuerpo Calloso: esta formado por fibras que ponen en relacion, son las correspondientes de la cortesa. La porcion posterior del cuerpo recibe el nombre de despleneo; su porcion central es el cuerpo, y su extremidad anterior, genu o rodilla, se continua como una delgada membrana, lamina rostral que se extiende, hasta la comisura anterior.
Piso del lll Ventrículo: El quiasma óptico señala el límite rostral del tercer ventrículo. Su porción anterior es concida como la eminencia media y de su vértice se origina el tallo hipofisiario. El flanco posterior de la eminencia media presenta un engrosamiento central, el túber cinéreo, formado por un conjunto de pequeños núcleos hipotalámicos.
Paredes en lll Ventrículo: Tálamo- Hipotálamo. La pared lateral del diencéfalo, está cruzada de delante a atrás por un surco poco profundo, el surco hipotalámo que la divide en dos porciones, una dorsal, el tálamo y una ventral, el hipotalámo.
Techo del lll Ventrículo: tela coroidea anterior: éste tercer ventrículo es una cavidad con piso, techo y paredes laterales.
Lámina terminal : es una delgada membrana que limita anteriormente la cavidad del tercer ventrículo, extendida entre el quiasma óptico y la comisura anterior. Por encima y por delante del quiasma, la lámina forma pequeña evaginación conocida como receso supraquiasmatico.
Tálamo
El tálamo es una gran masa ovoide de sustancia gris que forma la mayor parte del diencéfalo. Es una región de gran importancia funcional y sirve como estación celular para todos los sistemas sensitivos principales (excepto la vía olfatoria). El tálamo se sitúa a cada lado del tercer ventrículo .El extremo anterior del tálamo es estrecho y redondeado, y forma el limite posterior del agujero interventricular. El extremo posterior . se expande para formar el pulvinar, que sobresale horizontalmente por encima del colículo superior (tubérculo cuadrigémino superior) y el brazo conjuntival superior. El cuerpo geniculado lateral forma una pequeña elevación en la cara inferior de la porción lateral del pulvinar.
La cara superior del tálamo está cubierta intemamente por la tela coroidea y el fómix (trígono), y lateralmente por epéndimo, y forma parte del piso del ventrículo lateral; la parte lateral está oculta en forma parcial por el plexo coroideo del ventrículo lateral .
La cara inferior tiene continuidad con el tegmentum (calota) del mesencéfalo
La cara medial del tálamo forma la parte superior de la pared lateral del tercer ventrículo y habitualmente está conectada con el tálamo del lado opuesto por una banda de sustancia gris, la comisura gris intertalámica.
La cara lateral del tálamo está separada del núcleo lentiforme por la banda de sustancia blanca denominada cápsula interna
Subdivisiones del tálamo
El tálamo está cubierto en su cara superior por una delgada capa de sustancia blanca denominada estrato zonal y en su cara lateral por otra capa, la lámina medular externa. La sustancia gris del tálamo está dividida por una hoja vertical de sustancia blanca, la lámina medular interna, en una mitad medial y otra lateral. Por su parte anterosuperior, la lámina medular interna se desdobla en forma de Y. Así, el tálamo se subdivide en tres partes principales: la parte anterior se sitúa entre las ramas de la Y y la parte medial y la lateral se localizan a los lados del tronco de la Y.
Cada una de las tres partes del tálamo contiene un grupo de núcleos talámicos. Además, grupos nucleares más pequeños se encuentran en la lámina medular interna y algunos se hallan en las caras medial y lateral del tálamo. La descripción detallada de los núcleos
El tálamo es una estación celular muy importante y recibe los principales haces sensitivos (con excepción de la vía olfatoria). Se lo considera una estación en la que gran parte de la información se integra y retransmite a la corteza cerebral y muchas otras regiones subcorticales. También desempeña un papel clave en la integración de diversas funciones viscerales y somáticas.
Epitálamo
El epitálamo consiste en los núcleos habenulares y sus conexiones, y la glándula pineal.
La habénula es un pequeño grupo de células nerviosas de situación inmediatamente medial a la cara posterior del tálamo. Recibe fibras aferentes del núcleo amigdalino en el lóbulo temporal, a través de la estría medular del tálamo; otras fibras vienen de la formación del hipocampo a través del fórnix. Algunas de las fibras de la estría medular del tálamo atraviesan la línea media y llegan al núcleo de la habénula contralateral; estas últimas fibras forman la comisura habenular .Los axones del núcleo de la habénula se dirigen hacia el núcleo interpeduncular en el techo de la fosa interpeduncular, el techo del mesencéfalo, el tálamo y la formación reticular del mesencéfalo. Se cree que el núcleo de la habénula es un centro de integración de vías aferentes olfatoria, viscerales y somáticas.
El cuerpo o glándula pineal es una pequena estructura cónica unida al diencéfalo por el talío pineal. Se proyecta hacia atrás, de modo que su localización es posterior al mesencéfalo. La base del tallo pineal presenta un receso que tiene continuidad con la cavidad del tercer ventrículo. La parte superior de la base del tallo contiene la comisura habenular; la parte inferior de la base del tallo contiene la comisura posterior.
En el corte microscópico se observa que la glándula pineal está incompletamente dividida en lóbulos por tabiques de tejido conectivo que se extienden hacia su interior desde la cápsula. Se hallan dos tipos de células en la glándula, los pinealocitos y las células gliales. Progresivamente con la edad se acumulan concreciones de material calcificado denomi -nadas acérvulas o arenilla cerebral en la glándula pineal.
La glándula pineal no posee células nerviosas, pero recibe fibras simpáticas adrenérgicas procedentes de los ganglios simpáticos cervicales superiores, que corren en asociación con los vasos sanguíneos y los pinealoci
En la actualidad se considera que la glándula pineal, a la que alguna vez se atribuyó escasa importancia, es una glándula endocrina capaz de influir sobre las actividades de la hipófisis, los islotes de Langerhans, la paratiroides, las suprarrenales y las gónadas. Las secreciones pineales llegan a sus órganos blancos el torrente circulatorio o el líquido cefalorraquideo. Sus acciones son en su mayor parte inhibidoras; inhiben de manera directa la producción de hormonas o inhiben de manera indirecta la secreción de factores de liberación por parte del hipotálamo.
En experimentos realizados en animales se ha demostrado que la actividad pineal tiene un ritmo circadiano, influido por la luz. Se ha hallado que la glándula es más activa durante la oscuridad. La probable vía procedente de la re-tina sigue un trayecto hacia el núcleo supraquiasmático del hipotálamo, luego hacia la ca-lota del mesencéfalo, y después por el haz reticuloespinal a la eferencia simpática en la parte torácica de la médula espinal.
En la glándula pineal hay altas concentraciones de melatonina y serotonina. La liberación de noradrenalina de las terminaciones simpáticas de la glándula probablemente estimuía la liberación de estas sustancias por los pinealocitos.
Hipotálamo
El hipotálamo es la parte del diencéfalo que se extiende desde la región del quiasma óptico hasta el límite caudal de los cuerpos mamilares. Se encuentra por debajo del surco hipotalámico en la pared lateral del tercer ventrículo. El hipotálamo controla e integra las funciones del sistema nervioso autónomo y los sistemas endocrinos, y desempeña un papel vital en el mantenimiento de la homeostasis corporal. Interviene en actividades tales como la regulación de la temperatura corporal, los líquidos corporales, el hambre y la sed, la conducta sexual y las emociones.
Por delante del hipotálamo hay un área que se extiende hacia adelante desde el quiasma óptico hasta la lámina terminal y la comisura blanca anterior; se denomina área preóptica. Caudalmente, el hipotálamo se fusiona con la calota del mesencéfalo. Por encima del hipotálamo se halla el tálamo y por su parte inferolateral se encuentra la región subtalámica.
Cuando se observa desde abajo, se aprecia que el hipotálamo está relacionado con las siguientes estructuras, desde adelante hacia atrás: 1) el quiasma óptico, 2) el tuber cinereum y el infundíbulo y 3) los cuerpos mamilares El quiasma óptico es un haz aplanado de fibras nerviosas situado en la unión entre la pared anterior y el piso del tercer ventrículo. La cara superior está adherida a la lámina terminal y por abajo se reíaciona con la hipófisis, de la cual está separada por la tienda de la hipófisis (diaphragma sellae). Los ángulos anterolaterales del quiasma tienen continuidad con los nervios ópticos y los ángulos posterolaterales con las cintillas ópticas. En su cara superior se encuentra una pequeña cavidad, el receso óptico del tercer ventrículo.
El origen y destino de las fibras nerviosas del quiasma óptico se describen en la página 388. Es importante recordar que las fibras que se originan en la mitad nasal de cada re-tina atraviesan el plano medio en el quiasma e ingresan a la cintilla óptica del lado opuesto.
Estructura Anatómica
Si se daña una estructura anatómica cerebral debido a un error de posicionamiento, las consecuencias para el paciente pueden ser graves, pudiendo ocasionarle daños funcionales irreparables o, en el peor de los casos la muerte. Desde los orígenes de la neurocirugía se ha tratado de minimizar estos riesgos. Una de las herramientas de ayuda son los atlas cerebrales. Desde los más sencillos, atlas impresos, atlas digitalizados, hasta los más sofisticados, atlas deformables y probabilísticos.
En esta línea de investigación se han estudiado los distintos tipos de atlas. Se ha desarrollado un nuevo modelo de atlas deformable y en futuro próximo se pretende realizar su validación clínica.
HIPOFISIS: Ubicación. Relaciones. División. Irrigación. Sistema porta hipofisario.
Surco Hipotalàmico: es un surco profundo que se divide en dos porciones, una dorsal, el talàmo y una ventral, el hipotalàmo. Los dos lados se conectan,de manera incostante, en la lìnea media, por un puente de substancia gris o masa intermedia.
15. HIPOTÁLAMO
Limites:
En secuencia anteroposterior se pueden distinguir en él una región anterior, o supraoptica en relación con el quiasma óptico, una zona intermedia o tuberal, en relación con el infundíbulo, y una zona posterior o mamilar. En cortes transversales se consideran dos regiones, una medial, vecina a la pared del ventrículo donde se agrupan predominantemente los núcleos y una lateral por donde cursan las fibras que al él llegan y que de él salen.
Significado funcional:
El hipotálamo es el centro donde llega la información vísceras y es el ultimo eslabón del corredor neurovegetativo y donde se regula el funcionamiento viceral. Tiene funciones endocrinas por su relación con la adeno y la neurohipofisis y se relaciona con el sistema límbico, donde las sensaciones se convierten en emociones.
Núcleos:
Núcleo supraópticos: De neuronas grandes y redondeadas; se encuentran situadas por encima del tracto óptico cerca del quiasma.
Núcleo preóptico: Situado un poco por delante del anterior y entre este y la comisura anterior.
Núcleo paraventricular: Situado en la porción mas dorsal de esta región. Sus neuronas son similares a las del núcleo supraóptico.
Núcleos de la región tuberal: Las neuronas de esta región se distribuyen laxamente entre los núcleos dorsomedial, ventromedial, área hipotalámica dorsal y núcleos arciformes, que se continua por el piso del tercer ventrículo en el del lado opuesto.
Núcleos de la región mamilar: En la región mamilar están situados ventralmente los cuerpos mamilares, con sus núcleos lateral y medial y el área hipotalámica posterior situada dorsalmente en ellos.
Vías aferentes:
Fascículo cerebral medio.
Vía del fórnix.
Tálamo - hipotalámico.
Fascículo pálido - hipotalámico.
Estría terminal.
Pedúnculo mamilar.
Vías eferentes:
Fascículo mamiotalámico.
Conexiones con la hipófisis.
Fibras descendientes del hipotálamo.
Relaciones con el sistema neurovegetativo:
Las porciones anteriores del hipotálamo están en relación con el sistema parasimpático y su porción posterior con el simpático. Así. El estimulo de esta produce aumento de la presión sanguínea, midriasis e inhibición de la motilidad gastrointestinal y el estimulo de las porciones anteriores da, por contraste, signos de actividad parasimpático como vaso dilatación, miosis, disminución del ritmo cardiaco, aumento del ritmo respiratorio, defecación y micción.
Relaciones temperatura:
Este control se basa en el equilibrio de dos mecanismos, uno productor de calor y otro disipador de el. El primero tiende aumentar la temperatura a través del estimulo del metabolismo basal, de la vasoconstricción periférica y del aumento del ritmo cardiaco. El centro que regula estos fenómenos se encuentra en la región posterior del hipotálamo. El mecanismo de disipación del calor actúa a través de la vasodilatación, la sudoración y el aumento de la ventilación pulmonar y el centro de su regulación se encuentra en la parte anterior del hipotálamo. Entre la comisura anterior y el quiasma óptico.
Relaciones endocrinas:
Las conexiones directas del hipotálamo con el lóbulo posterior de la hipófisis ponen de manifiesto su importancia en la producción de la hormona antidiurética. La sección del fascículo hipotálamo hipofisiario da como resultado la aparición de la diabetes insípida.
Relación con el apetito:
Lesiones experimentales que comprometen los núcleos ventromediales del hipotálamo, dan como resultado animales con voracidad insaciable; por el contrario, lesiones en el área hipotalámica lateral produce marcada perdida del apetito y enflaquecimiento. El funcionamiento de estos centros es regulado por los nivel de glucosa sanguínea.
TALAMO
Forma:
Presenta forma ovoidea cuyo diámetro mayor anteroposterior alcanza 25 mm, su ancho es de 20 mm y su altura 25 mm.
Cubiertas:
La superficie dorsal esta recubierta por una delgada lamina de substancia blanca, el stratum zonale, y hace parte del piso del ventrículo lateral.
La cara lateral esta separada de las fibras de la cápsula interna por la lamina medular externa.
La superficie medial forma parte de la pared del III ventrículo y en su parte media se une por un puente de substancia gris conocida como masa intermedia.
División:
El tálamo esta divido en dos porciones, lateral y medial, por una lamina de substancia blanca, la lamina medular interna.
Significado funcional:
Es un centro de integración de gran importancia que recibe las señales sensoriales y donde las señales motoras de salida pasan hacia y desde la corteza cerebral.
Localización y función de los núcleos:
Núcleos de la línea media: Son un grupo de células que se encuentran por debajo de la pared media del tálamo que se proyectan sus fibras a otros núcleos del tálamo y al hipotálamo.
Núcleo dorsomediano: presenta una porción medial por células que reciben fibras de los núcleos de la línea media para luego proyectarlas luego al hipotálamo.
Núcleos dorsal y posterior: reciben fibras de otros núcleos del tálamo para luego proyectarlas hacia la corteza temporal y parietal.
Núcleo de la lamina medular media: hacen conexiones con el fascículo espinotalámico con el cuerpo estriado y con la corteza.
Núcleo ventral anterior: recibe fibras del globos pálidos a través del fascículo talámico.
Núcleo ventral lateral: recibe fibras de la braquioconjuntiva y del núcleo rojo para luego ser proyectadas a la corteza motora. A través de estas vías se produce una comunicación entre el cerebelo y cerebro.
Núcleo ventral posterior: presenta uno a su vez una parte lateral. Recibe fibras del lemnisco medio y fibras espirotalámicas.
Núcleo pulvinar: Corresponde al polo posterior del tálamo, es voluminoso, recibe fibra de otros núcleos del tálamo y luego las proyecta a la corteza occipital y temporal.
9. Núcleo reticular: esta formado por pequeñas células nerviosas, q ue hacen conexión con los fascículos tálamocorticales y corticotalámicas. Son células llamadas inhibidoras que intervienen en la atención selectiva.
17. VÍAS ÓPTICAS
Componentes a nivel de la retina:
La retina esta constituida en esencia por una cadena de tres neuronas; la mas periférica de ellas tiene terminaciones grandemente diferenciadas (conos y bastones) para responder a los estímulos luminosos. La segunda neurona es la neurona bipolar que recibe los estímulos de conos y bastones, y establece sinapsis con la neurona ganglionar.
Igualmente está formada por diez capas entre las que destacan el epitelio pigmentario, la capa más externa, y la capa sensorial, compuesta por fotorreceptores.
Para poder observar la retina, dada su localización, es preciso usar un oftalmoscopio, en el proceso denominado estudio del fondo de ojo. En esta visualización de la retina sirven de referencia dos estructuras muy importantes: la papila y la mácula, que deben ser examinadas siempre al realizar un estudio del fondo de ojo. La papila corresponde al punto en que el nervio óptico se continúa con la retina, mientras que la mácula representa el punto más sensible de toda la retina, es decir, la zona de máxima visión.
Nervio óptico:
Es en realidad un fascículo cerebral. Sus fibras, a partir de la zona cribosa de la esclerótica, están recubiertas por una vaina de mielina no formada por células de Schwan, como en los nervios periféricos, sino por células de oligodendroglia.
Quiasma óptico:
El nervio óptico al entrar al endocraneo y en el surco óptico (quiasma), las dos terceras partes de sus fibras que corresponden a los nasales de la retina se entrecruzan con los del lado opuesto formando el quiasma óptico.
Tracto óptico:
Una tercera parte de las fibras procedentes de la porción lateral de la retina se cruzan. Después del quiasma estas fibras ópticas toman el nombre de TRACTO o ANTILLA ÓPTICA, las que se dirigen al cuerpo geniculado lateral del tálamo.
Campo visual:
Es el espacio captado por los ojos con la mirada fija en un punto, el campo visual recibe una denominación para cada ojo, considerándose en él dos mitades, una nasal y otra temporal, separadas por un eje vertical y dos mitades, superior e inferior.
Anopsia:
Perdida de visión.
Hemianopsia:
Perdida de la visión en medio campo visual, nasal y temporal respectivamente, ocurre cuando hay lesión del tracto óptico.
Reflejos normales:
Reflejo a la luz: Al iluminarse la retina produce la contracción del músculo constrictor de la pupila. Puede igualmente notarse que esta respuesta es compartida por ambas pupilas, aunque el estimulo sea unilateral; lo que se llama respuesta o reflejo consensual.
Reflejo de la acomodación: Este reflejo controla la contracción del músculo ciliar de la cual dependen el grado de convexidad del cristalino. Cuando los rayos llegan divergentes, dicha convexidad debe aumentarse para producir la misma convergencia en el mismo sitio.
18. VÍAS OLFATORIAS
Mucosa:
El órgano del olfato esta formado por una pequeña zona de la mucosa pituitaria, que alcanza unos 5 cm2 y comprende el techo de las fosas nasales, parte de los cornetes superiores y parte del tabique nasal. En esta área se encuentran las células olfatorias, que son neuronas bipolares cuya rama periférica que llega hasta la superficie de la mucosa y se pone en contacto con las partículas odoríficas y cuya rama central atraviesa la lamina cribosa del etmoides para llegar al bulbo olfatorio.
Nervio olfatorio:
Se origina en el bulbo olfatorio pasando por la lamina cribosa.
Bulbo:
Esta alojado en la parte anterior del surco olfatorio en la base del lóbulo frontal; in situ queda sobre puesto a la lamina cribosa del etmoides, a través de la cual le llegan los filetes del nervio olfatorio o primer par craneal.
Cintilla y trígono olfatorio:
El bulbo se continua con la cintilla olfatoria, la cual a nivel del espacio perforado anterior forma un pequeño ensanchamiento triangular o Trígono olfatorio, cuyos ángulos posteriores se originan las estrías olfatorias lateral y medial.
Tubérculo:
Ocupa la parte anterior del espacio perforado anterior; se observa en algunos mamíferos y ocasionalmente en el hombre como una pequeña prominencia formada por neuronas que reciben la sinapsis de fibras de la cintilla olfatoria.
Banda diagonal de Broca:
En la parte posterior del área perforada anterior y extendiéndose entre la porción de la corteza, ventralmente situada en la rodilla del cuerpo calloso y la región perimigdalina se encuentra una prominencia, no muy aparente, por lo general recibe el nombre de BANDA DIAGONAL DE BROCA.
Núcleos:
Núcleo olfatorio anterior.
Núcleo del tubérculo olfatorio.
Núcleo olfatorio de la estría lateral.
Conexiones:
Bulbo olfatorio y sus conexiones: El bulbo olfatorio es una porción derivada en el embrión de la porción basal de los hemisferios cerebrales; en un principio muestra en su centro una cavidad que se comunica con los ventrículos laterales, pero pronto desaparece.
19. SISTEMA LIMBICO
- PORCIONES DEL CEREBRO QUE LO FORMAN:
Está formado por porciones del cerebro de precoz desarrollo filogénico y ontogénico, situadas en la porción mesial del hemisferio.
Por sus directas conexiones con estructuras olfatorias, inicialmente se relacionó el lóbulo límbico con la olfacción y por ello recibió también el nombre de rinencefalo.
En la actualidad se sabe que en él asientan funciones de mayor importancia relacionadas con el mantenimiento o supervivencia del individuo o de la especie y con la integración emocional del comportamiento.
-
Porciones del lóbulo límbico:
Está formado el lóbulo límbico por las estructuras que conforman del hipocampo, la corteza parahipocampal y el núcleo amigdaloide y sus conexiones.
-
LOCALIZACIÓN DEL HIPOCAMPO:
Es una porción del archipallium invaginada en el interior de la prolongación temporal del ventrículo lateral de la cual forma su piso; en su parte anterior termina en forma de tres o cuatro engrosamientos o digitaciones del hipocampo.
-
CORTEZA:
Giro dentado o fascia dentada, es una delgada banda de corteza situada medialmente al asta de Amón y delimita, en la parte interna del lóbulo temporal, la fisura del hipocampo. Debe su nombre a una serie de surcos transversales que muestra en su superficie. En el desarrollo inicial de la corteza en el embrión hemos visto diferenciar el archicortex como una banda de corteza situada dorsalmente al tálamo. Con la subsiguiente remodelación del hemisferio, el archicortex alcanza su mayor desarrollo en el lóbulo temporal (hipocampo).
La corteza parahipocampal es la que continúa medialmente el giro dentado sobre el giro del hipocampo; constituye una transición entre el archicortex y el neocortex del lóbulo temporal y citoarquitectónicamente constituye la corteza 28 de Brodman.
-
NÚCLEO
Es en realidad un complejo de varias masas nucleares que se agrupan en la porción del Uncus del lóbulo temporal y que se pueden dividir en dos grupos principales, el grupo Corticomedial, y el basolateral.
El núcleo amigdaloide recibe conexiones de la corteza parahipocampal, del tubo olfatorio y de la porción basal del lóbulo frontal a través del fascículo uncinado; tiene también conexiones con el hipocampo y el giro dentado y con el área septal a través de la estría terminal.
-
RELACION COMPORTAMIENTO O CONDUCTA:
Es el conjunto de manifestaciones o actuaciones expresas por el individuo, aparentes para un observador. Desde un punto de vista experimental, tanto por destrucción como por estímulo de algunas de sus estructuras, se ha demostrado que el sistema límbico representa un centro de coordinación de este tipo de comportamiento. Además de los fenómenos externos, la conducta se acompaña de un componente interno, emotivo, referible como ¨tono afectivo, que puede ser de agrado, desagrado, de placer, de incomodidad o de atención, etc., y que puede ser a veces reconocible en la expresión del individuo y en sus manifestaciones viscerales. También la coordinación de este componente afectivo como las manifestaciones orgánicas se lleva a efecto a través de las conexiones neuronales del sistema límbico.
-
TEMOR :
Aprehensiones e inhibiciones naturales en el animal pueden desaparecer por lesión de la corteza medial del lóbulo temporal. Experimentalmente se puede comprobar en micos, que normalmente manifiestan pánico ante las serpientes, como las toman sin temor y aun las llevan a su boca con la mayor confianza, luego de una lesión de este tipo.
-
IRA :
Hemos visto que por estímulo de ciertas zonas del hipotálamo se puede desencadenar una reacción de ira intensa en el animal. Es conocido también el hecho que el neocortex ejerce una acción represiva sobre esta reacción, ya que animales decorticados la manifiestan fácilmente con estímulos mínimos.
-
COMPORTAMIENTO SEXUAL:
En gran parte la actividad sexual del animal es debida a la tasa y tipo de hormonas sexuales en la sangre circulante. En el sistema nervioso existen receptores que al ser estimulados por ellas desencadenan los circuitos coordinados de tales acciones.
-
COMPORTAMIENTO ALIMENTICIO:
Lesiones en los núcleos amigdaloides producen cambios en los hábitos alimenticios como hiperfagia o ingesta indiscriminada de toda clase de alimentos. Debemos recordar que cambios similares pueden observarse por lesiones en los núcleos mediales del hipotálamo, lo que indica la directa relación funcional entre ambas estructuras.
20. LA CORTEZA CEREBRAL
- LOCALIZACIÓN:
La corteza cerebral es una extensa capa de substancia gris que recubre superficialmente los hemisferios cerebrales. Más de la mitad de su extensión se encuentra escondida en la profundidad de surcos y fisuras.
Está constituida por un agregado de neuronas y fibras aferentes, dispuestas de una manera altamente ordenada, por el tejido glial característico de los centros nerviosos y por una riquísima red capilar.
- CAPAS:
-
Capa molecular o plexiforme.
-
Capa granular externa o capa de las pirámides pequeñas.
-
Capa de células piramidades.
-
Capa granular interna.
-
Capa ganglionar o de las grandes pirámides.
-
Capa de células fusiforme o polimorfas.
- CONEXIONES AFERENTES, EFERENTES:
-
AFERENTES :
A la corteza llegan fibras de diverso origen; en primer lugar las que ya hemos descrito procedentes de los núcleos talámicos que constituyen las fibras aferentes específicas, con una distribución conocida en las distintas áreas corticales.
En segundo lugar las fibras de asociación que ponen en relación varias zonas de la corteza, y por último, un grupo de fibras que establecen conexiones directas con la formación reticular del tallo cerebral y que ha recibido el nombre de sistema de proyección difuso.
-
EFERENTES :
Los axones de las células piramidales pequeñas y granulares de las capas 1 a 4 se ramifican y terminan de la corteza misma. Los de las neuronas mayores, en especial aquellas de las capas 5 y 6, penetran a la substancia blanca y ahí siguen diversos caminos. Unas son fibras de asociación que ponen en relación varias partes del mismo hemisferio; otras son fibras comisurales que pasan al otro hemisferio a través del cuerpo calloso y terminan en sitios homologables al de su origen y, por último, las fibras de proyección que van a hacer sinapsis con otros núcleos celulares, como el tálamo.
21. REPRESENTACIONES CORTICALES:
-
ENUMERARLAS :
Representación motora de la corteza.
Representación de movimientos.
Representación bilateral de movimientos.
Representación sensorial rolándica.
Representación vestibular.
Representación del gusto.
Representación neurovegetativa de la corteza.
-
AREA MOTORA:
Las distintas partes de éste se encuentran representadas de una manera ordenada de arriba hacia abajo a lo largo de la región precentral con la siguiente secuencia: en la parte inicial de esta región, en la superficie medial del hemisferio, se encuentra la representación motora del pie y pierna; a continuación y ya sobre la superficie dorsolateral se encuentra la representación de los músculos del muslo; luego los de la cadera; los del tronco; los de los hombros; los del brazo; mano en general y luego los movimientos propios de cada dedo meñique, anular, medio, índice y pulgar en orden descendente.
-
AREA DE REPRESENTACIÓN CORTICAL:
El estudio topográfico de las funciones de la corteza cerebral se inició a mediados del siglo pasado con las observaciones de Broca en 1861 quien correlacionó la afasia que presentaban algunos pacientes con lesiones de la base de la tercera circunvolución frontal y con las observaciones de jackson en 1864 acerca de la existencia de lesiones en la región precental en pacientes que presentaban convulsiones epilépticas. Estos trabajos iniciales y los que a ellos siguieron, cuidadosamente elaborados a través del último siglo, nos han dado un cuadro completo de las representaciones funcionales y sus áreas respectivas en la corteza del cerebro humano.
-
AREA PREMOTORA:
El área es caracterizada por la existencia de las grandes pirámides de Betz en la capa V de la corteza, está restringida en el hombre a la profundidad y flanco anterior de la fisura central; las respuestas motoras se consiguen en esta zona con una intensidad mínima de estimulación eléctrica.
22. SISTEMA ESTRAPIRAMIDAL:
La actividad motora somática del organismo tiene una amplia ordenación jerárquica en los distintos niveles del sistema nervioso; movimientos más sencillos, como el reflejo de extensión, tienen los elementos anatómicos para su ejecución limitados a un solo segmento medular; otros, que tienen que ver con el equilibrio y las reacciones posturales, dependen de los impulsos nacidos en el vestíbulo y por lo tanto son influenciados por los núcleos y vías vestibulares. Los principales componentes de este sistema son el cuerpo estriado, el globus pallidus, el núcleo subtalámico, la zona incerta, el núcleo prerrubral, el núcleo rojo y la substancia nigra; La principal vía eferente de este sistema, a través de la cual sus impulsos se ejercen sobre las neuronas medulares, la constituyen los fascículos reticuloespinales.
-
ENUMERAR LOS NÚCLEOS DE LA BASE:
El cuerpo estriado.
El núcleo subtalámico.
la zona incerta.
El núcleo prerrubral.
La substancia nigra.
El núcleo rojo.
-
CUERPO ESTRIADO:
Recibe fibras de la corteza de los núcleos intralaminares del tálamo y de la substancia nigra. Sus conexiones eferentes se hacen sobre todo con el globus pallidus y la substancia nigra. El globus pallidus aparece como centro de recepción y distribución de los impulsos de los núcleos básales. Recibe impulsos del putamen y el núcleo subtalámico y sus fibras eferentes forman tres fascículos, el ANSA lenticularis, el fascículo lenticular y el fascículo subtalámico.
-
REGION SUBTALAMICA:
Es una masa gris, ovoidea, situada ventralmente al tálamo y en su porción posterior. Está separado el tálamo por el fascículo lenticular y la zona incerta. Sus principales conexiones se establecen con el globus pallidus a través del fascículo subtalámico.
-
SIGNIFICADO FUNCIONAL:
La función precisa de los distintos núcleos extrapiramidales en el complejo control de la actividad motora dista mucho de estar completamente definida, en especial por los pocos resultados que arroja el análisis experimental de sus estímulos o destrucción y ha sido por la correlación de los casos de patología humana como se han logrado definir algunas de sus acciones.
-
TONO MUSCULAR:
Una de las principales manifestaciones de las lesiones de los núcleos básales en el hombre es el aumento del tono muscular. Recordemos que la rama aferente del reflejo que determina la contracción tónica del músculo es la fibra anuloespinal originada en el huso neuromuscular, y que ella puede ser estimulada tanto por la tracción del músculo como por la contracción de la fibra intrafusal.
-
COORDINACIÓN :
El sistema extrapiramidal tiene una acción directa sobre el sistema voluntario. Por sus conexiones con el tálamo influye sobre la actividad funcional de la corteza y por sus conexiones reticulares actúa sobre la actividad de las neuronas y de los núcleos del tallo cerebral y de la médula espinal. Su intervención se relaciona sobre todo con la adecuación del tono muscular necesario para un movimiento determinado, en su correlación temporal, en la coordinación del juego de músculos protagonistas y antagonistas y en la producción de movimientos asociados para la correcta ejecución de un movimiento voluntario. Un ejemplo de estas acciones lo podemos notar en la ejecución de un pequeño movimiento de los dedos en la escritura; vemos que en esta acción se requiere la fijación del hombro, del brazo y del antebrazo; la adecuación del tono de los músculos flexores y extensores de la muñeca y la coordinación de los movimientos de los músculos flexores y extensores de la muñeca y la coordinación de los movimientos d los músculos flexores y extensores de los dedos. Todas estas acciones requieren evidentemente de un amplio control del cual no somos conscientes.
-
CORRELACIÓN CLINICA:
Los síndromes extrapiramidales en el hombre pueden ser considerados en dos categorías según el predominio de determinadas manifestaciones: en una predominan el aumento del tono muscular y el temblor, características del parkinsonismo, y en la otra dominan la producción de movimientos anormales, como en las coreas, la atetosis y el hemibalismo.
23. FORMACION RETICULAR
La formación reticular corresponde a una amplia zona central del tallo cerebral formada por agregados nucleares no muy compactos en su organización, que reciben conexiones de muy distinto origen y que envían sus fibras tanto en sentido ascendente, hasta el diencéfalo, como desde dentro hasta la médula espinal. Desde el punto de vista funcional, constituye un centro de coordinación sensitivo y motor de naturaleza inespecífica ya que integra estímulos de muy diverso origen y mantiene un estado de actividad básico e indiferenciado en distintos niveles del sistema nervioso.
Núcleos de la Formación Reticular.
En la mitad inferior del bulbo se aprecian los núcleos reticular central y reticular lateral. En la mitad superior, dorsalmente a la oliva bulbar, y en la zona vecina de la protuberancia se localiza medialmente en núcleo reticular magnocelular, formado por grandes neuronas de tipo motor y lateralmente el núcleo parvocelular con neuronas de pequeño tamaño. En el tercio medio de la protuberancia esta el núcleo reticular caudal del puente (reticularis pontis caudalis) y en su tercio superior el reticular oral del puente ( o reticularis pontis oralis). En el mecencéfalo, las células reticulares se dividen en dos áreas principales, en núcleo cuneiforme, dorsalmente, y el subconeiforme en la región lateral del segmento.
CONEXIONES DE LA FORMACION RETICULAR.
Conexiones con la Médula Espinal. Las vías reticulares se encuentran tanto en los cordones anteriores como en los laterales. Torvic y Brodal han estudiado su origen en el gato y lo han señalado en el bulbo y en el puente, sobre todo en los núcleos magnocelular, caudal del puente y región inferior del núcleo pral. Según los mismos autores, las fibras originadas en la protuberancia descienden por la parte media del cordón anterior de la médula, por el mismo lado de su origen, mientras que las bulbares ocupan los cordones laterales y son tanto directas como cruzadas.
Las fibras espinorreticulares tiene su origen en células de distintos niveles medulares y terminan difusamente en todos los núcleos reticulares. Además ellos reciben colaterales de otros fascículos ascendentes, en especial los espinotalámicos.
Sistema reticular ascendente. Se conoce con este nombre el conjunto de fibras que ascienden de los núcleos reticulares desde el bulbo hasta el diencéfalo, y que hacen parte del fascículo tegmental central. En el límite del diencéfalo con el mesencéfalo unas de las fibras pasan al tálamo y terminan en los núcleos intralaminares, mientras que otras, sobre todo las originadas en el tegmento mesencefálico, van al hipotálamo, unas como parte del pedúnculo mamilar y otras como parte del sistema periventricular que rodea el tercer ventrículo para llegar a los distintos núcleos hipotalámicos y a los del área sectal. Por medio de las concesiones de los núcleos intralaminares con los núcleos talámicos el sistema reticular ascendente ejerce una amplia acción sobre todas las áreas de la corteza cerebral, manifiesta tanto en cambios del estado de conciencia como de la actividad eléctrica cortical.
Concesiones corticorreticulares. Se originan en distintas áreas de la corteza pero principalmente en las regiones centrales sensitivas y motoras. Se distribuyen bilateralmente y terminan sobre todo en los núcleos del bulbo y del puente.
Conexiones con el cerebelo. Son tanto aferentes como eferentes. Las fibras reticulocerebelosas se originan en especial en los núcleos lateral y paramediano del bulbo y en el núcleo tegmento póntico de la protuberancia.
Otras conexiones de los núcleos reticulares. Tiene además la formación reticular concesiones con los sistemas sensoriales, como el sistema visual, a través de sus concesiones con el colículo superior, con los núcleos cocleares y vestibulares y con los núcleos del sistema extrapirámidal.
Nota funcional.
Si desde el punto de vista anatómico la formación reticular tiene conexiones con todos los niveles del sistema nervioso, también desde el punto de vista funcional ejerce sus influjo sobre las actividades motoras y sensitivas, somáticas y viscerales, y muy especialmente sobre la actividad cortical, manifiesta en la regulación de los estos de conciencia.
LA FORMACION RETICULAR Y LA ACTIVIDAD MOTORA.
Una primera indicación de la acción de la formación reticular sobre la actividad muscular es la llamada rigidez de descerebración secundaria a la sección del tallo cerebral a nivel del mesencefalo, la cual se ha explicado como una manifestación de la actividad excitadora reticular sobre los reflejos medulares. Por estimulación de la formación reticular del bulbo se puede producir inhibición tanto de la rigidez de descerebración como de la actividad muscular refleja y de la producida por estímulos corticales. Los estímulos en sitios más rostrales a esta zona inhibitoria bulbar tiene un efecto opuesto, es decir, facilitador de la actividad muscular, lo cual permitió delimitar zonas excitadoras u zonas inhibidoras dentro de la formación reticular. Para la ejecución normal de los movimientos las dos acciones se manifiestan de manera recíproca, es decir, que coordinan la acción de los músculos agonistas y antagonistas de acuerdo con el movimiento que se ejecuta según el principio de la inervación recíproca.
Las acciones reticulares sobre la actividad muscular se ejercen sobre la neurona alfa, bien sea por sinapsis directa o indirectamente a través de neuronas intercalares. Pero también se manifiesta a través del centro de la neurona gamma, es decir modificando la actividad del reflejo miotático originado en el huso neuromuscular.
Mencionemos por último que a través de sus concesiones con los núcleos reticulares, como el cerebelo y los núcleos del sistema estrapiramidal manifiestan su acción sobre la actividad muscular.
LA FORMACIÓN RETICULAR Y LA ACTIVIDAD ENDOCRINA.
El sistema endocrino actúa con mecanismos de autorregulación a través de los receptores hormonales hipotalámicos y la producción del principios de liberación de las distintas hormonas hipoficiarias, pero para la liberación de ciertas hormonas en especial de ACTH, son de gran importancia los estímulos nerviosos ascendentes, canalizados a través de las conexiones de la formación reticular con el hipotálamo.
El control nervioso de la secreción de ACTH fue claramente establecido con los trabajos de Selye sobre la acción de alarma “Stress”, para la superación de la cual el organismo requiere un aumento de la producción de esteroides corticosuprarrenales. Se ha demostrado experimentalmente que distintos estímulos nerviosos originados por la situación de emergencia ascienden por el tallo cerebral y de la porción rostral del mesencefalo pasan al hipotálamo, bien sea directamente a través del péndulo mamilar o indirectamente, vía fórnix y estría terminalis, después de haber alcanzado distintas zonas del rinencéfalo.
Se ha visto también que la adrenalina circulante estimula la liberación de ACTH a través del estímulo de los núcleos reticulares del mesencéfalo. A este respecto Porter demostró que las conexiones del mesencéfalo con el hipotálamo suprimen la acción de la adrenalina, lo que señala un eslabón adrenergético en la cadena neuronal responsable de dicho control.
LA FORMACION RETICULAR Y EL SISTEMA NERVIOSO AUTONOMO (SNA).
Células de la formación reticular del tallo cerebral, en especial del bulbo raquídeo, conforman los distintos centros de coordinación visceral que regulan el ciclo respiratorio y la actividad cardiaca y bascular de las cuales ya hablamos como los centros respiratorios, vasomotor entra otros.
24.MENINGES Y LIQUIDO CEFALORRAQUIDEO
Una meninge se refiere a cada una de las membranas de tejido conjuntivo que recubren el sistema nervioso central, formado por el encéfalo (telencéfalo, diencéfalo, mesencéfalo, protuberancia, bulbo raquídeo) y la médula espinal.
Las 3 meninges, duramadre (paquimeninge, externa), aracnoides y piamadre (leptomeninges), limitan 3 espacios, el epidural (entre el estuche osteoligamentoso —cráneo o raquis— y la duramadre), el subdural y el subaracnoideo. El espacio epidural es aprovechado en la médula para inyectar anestésicos locales consiguiendo anestesia temporal del abdomen y miembros inferiores.
Éstos se nombran de fuera hacia adentro: la Duramadre, el Aracnoides, y la Piamadre.
La Duramadre
La Duramadre: es una membrana inelástica gruesa y densa. La porción que incluye el cerebro diferencia en varios detalles esenciales de la que rodee los espinales de la médula, y por lo tanto él es necesaria describirlos por separado; pero en el mismo tiempo debe ser entendido distintamente que la membrana completa de dos formas una, y es continuo con uno a en foramen magnum.
La Duramadre Craneal ( encephali del mater del dura; El dura del cerebro) alinea el interior del cráneo, y sirve el propósito doble de un periostio interno a los huesos, y una membrana para la protección del cerebro. Se compone de dos capas, un interno o meníngeo y un externo o endosteal, conectado de cerca junto, excepto en ciertas situaciones, donde, como descrito, se separan para formar los senos para el paso de la sangre venosa. Su superficie externa es áspera y dura, y adhiere de cerca a las superficies internas de los huesos, las adherencias que son las más marcadas enfrente de las suturas y en la base del cráneo su superficie interna son lisas y alineadas por una capa de endotelio. Envía interior cuatro procesos que dividan la cavidad del cráneo en una serie de compartimentos libremente el comunicarse y la protección de las diversas partes del cerebro; y se prolonga a la superficie externa del cráneo, con el vario foramina que existen en la base, y llega a ser así continuo con el pericraneo; su capa fibrosa forma las envolturas para los nervios que pasan a través de estas aberturas. Alrededor del margen del foramen el magnum que es de 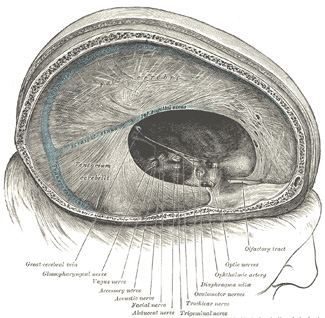
cerca adherente al hueso, y es continuo con la duramadre espinal.
F IG. 765- Duramadre y sus procesos expuestos quitando la parte de la mitad derecha del cráneo y del cerebro.
Procesos. -- los procesos de la Duramadre, que proyecta en la cavidad del cráneo, son formados por reduplicación de la capa interna o meníngea de la membrana.
La Hoz del cerebro
Nombrado tan de su hoz-como forma, es un proceso fuerte, arqueado que desciende verticalmente en la grieta longitudinal entre los hemisferios cerebrales. Es estrecho en el frente, donde se une a la crista galli del etmoides; y amplio detrás, donde está conectada con la superficie superior de la tiende del cerebro (cerebelli del tentorium). Su margen superior es convexo, y unido a la superficie interna del cráneo en la línea media, tan lejos parte posterior como la protuberancia occipital interna; contiene el sino sagital superior. Su margen más bajo es libre y cóncavo, y contiene el sino sagital inferior.
La Tienda del Cerebro (El cerebelli del tentorium) es una lamina arqueada, elevada en el centro, y la inclinación hacia abajo hacia la circunferencia. Cubre la superficie superior del cerebelo, y apoya los lóbulos occipitales del cerebro. Su frontera anterior es libre y cóncava, y limita una abertura oval grande, la incisura de la tienda, para la transmisión de los pendículos cerebrales. Es unida, detrás, por su frontera convexa, a los cantos transversales sobre la superficie interna del hueso occipital, e incluye el sino transversal; en frente, al ángulo superior de la pieza longitudinal del hueso temporal de cualquier lado, incluyendo los senos longitudinales superiores. En el ápice de la pieza longitudinal del hueso temporal las fronteras libres y unidas satisfacen, y, cruzando uno otro, se continúan adelante siendo fijadas a los procesos anteriores y posteriores del clinoideo respectivamente. A la línea media de su superficie superior la frontera posterior de la hoz se une, el seno recto que es colocado en su línea de la ensambladura.
F IG. 766- vista superior de la tienda.
El orificio de la tienda es un proceso triangular pequeño de la duramadre recibido en la muesca cerebelosa posterior. Su base se une, arriba, a la parte inferior y trasera del tentorium; su margen posterior, a la división más baja de la cresta vertical en la superficie interna del hueso occipital. Mientras que desciende, se divide a veces en dos dobleces más pequeños, que se pierden en los lados del foramen magnum.
Estructura. La duramadre consiste en el tejido fino fibroso blanco y por las fibras elásticos dispuestas en laminas aplanadas que son separadas imperfecto los espacios lacunar y los bloodvessels en dos capas, endosteal y meníngeo. La capa endosteal es el periostio interno para los huesos craneales, y contiene los bloodvessels para su fuente. En el margen del foramen el magnum que es continuo con el periostio que alinea el canal vertebral. La capa meníngea o de soporte es alineada en su superficie interna por una capa de mesotélio aplanado nuclear, similar a ésa encontrada en las membranas serosas.
Las arterias de la duramadre son muy numerosas. En la fosa anterior son las ramas meníngeas anteriores de la carótida etmoidal e interna anterior y posterior, y de una rama del centro meníngea. En la fosa media son las meníngeas medias y accesorias del maxilar interno; una rama del piramidal ascendente, que entra en el cráneo con foramen lacerum; ramas de la rama carótida, y recurrente interno del lagrimal. En la fosa posterior son ramas meníngeas del occipital, uno que entra en el cráneo con el yugular foramen, y otro a través del mastoideo foramen; el trasero meníngeo del vertebral; los ramas meníngeos ocasionales del piramidal ascendente, entrando en el cráneo con el yugular foramen y canal hipogloso; y una rama del centro meníngeo.
Las venas. Muchas de las venas meníngeas no se abren directamente en los senos, sino indirectamente con una serie de ampollas, llamadas lagunas venosas. Éstas se encuentran de cualquier lado del seno sagital superior, especialmente cerca de su porción central, y son a menudo por granulaciones aracnoides; también existen cerca de los senos transversales y rectos. Se comunican con las venas cerebrales subyacentes, y también con las venas diploicas y emisarias.
Los nervios de la duramadre son filamentos del ganglio semilunar, el el oftálmico, maxilar, de la mandíbula, del vago, y de los nervios hipoglosal, y del comprensivo.
LA ARACNOIDES
La aracnoides craneal, como la medular es delgada y avascular y se encuentra adherida a la superficie interna de la duramadre. Envía finas trabéculas que atraviesan el espacio subaracnoideo para fijarse en la píamadre y en algunos sitios, sobre todo en contacto con el seno longitudinal superior, forma vegetaciones de pequeñas papilas o vellosidades a nivel de las cuales el líquido cefalorraquídeo pasa a la circulación venosa. En el adulto muchas de estas vellosidades se calcifican y reciben el nombre de granulaciones de Pachioni.
LA PIAMADRE.
Recubre directamente la superficie cerebral, introduciéndose por surcos y fisuras; en ellas se ramifican las arterias corticales como un fino plexo arteriolar. Igualmente en ella se forman las venas corticales que va a drenar a los distintos senos de la duramadre.
Espacio subaracnoideo. Cisternas craneales. Es el espacio subaranoideo craneal de amplitud variable en sus distintas regiones debido a las irregularidades en la forma del encéfalo. En unos sitios es amplio y las cavidades resultantes reciben el nombre de Cisternas, las principales son:
Cisterna magna o cerebelomedular. Es la mayor de las cisternas y está limitada por el bulbo y el cerebelo. A ella se puede llegar por función de la dura a través del intersticio entre el atlas y el occipital, zona en la cual alcanza una profundidad de 12mm.
Cisterna superior. Queda limitada por el esplenio del cuerpo calloso y el vermis del cerebelo.
Cisternas de la base. Son espacios relacionados con la porción ventral de la protuberancia (cisterna póntica), la fosa interpeduncular (cisterna interpeduncular) y el quiasma óptico (cisterna quiestmática), entre los cuales existe una directa continuidad.
Cisterna lateral o silviana. Representa un delgado conducto correspondiente a la fisura lateral del cerebro.
LIQUIDO CEFALORRAQUIDEO
El Liquido Cefaloraquideo, (l.c.r.), es un liquido acuoso, incoloro como agua de roca, que normalmente llena los ventrículos cerebrales y los espacios subaracnoideos craneales y espinales. Su volumen se calcula entre 100 y 150 cc. Y de estos aproximadamente la cuarta parte se encuentra en las cavidades ventriculares. Tiene una densidad de 1005 - 1008; su pH es de 7.4 y su presión osmotica es superior a la del plasma.
COMPOSICIÓN DEL LÍQUIDO CEFALORRAQUIDEO (mg/100 ml)
Sodio 325
Cloruro 445
Potasio 11
Calcio 5
Magnesio 2.5
Bicarbonato 65
Proteínas 20 - 30
Glucosa 60 - 70
Urea 20
Creatinina 1.2
El líquido cefalorraquideo representa un sistema de defensa del sistema nervioso central, ya que forma una capa líquida protectora entre éste y la pared ósea craneovertebral, que amortigua los golpes y compresiones que fácilmente lo afectarían. En el cráneo sirve además como medio de flotación del cerebro, y disminuye así su peso real de unos 1400g a unos 50g de su peso efectivo, con lo cual los riesgos de daño con los desplazamientos se disminuyen aún más.
Circulación del l.c.r. la mayor cantidad del l.c.r. se forma entre los plexos coroides de los ventrículos cerebrales desde donde pasa a los espacios subaracnoideos de la cisterna magna a través de los orificios laterales (de luschka) y medial (de magendie) del cuarto ventrículo. Del espacio subracnoideo es drenado a los senos dúrales por las vellosidades subaranoideas y en menor porción por los linfáticos de la dura a través de los fondos del saco aracnoideo de los nervios raquídeos y craneales.
Las vellosidades aracnoideas son pequeñas proyecciones de las aracnoides que perforan la pared de los senos dúrales; en su interior presentan estrachos canales de pocas micras de diámetro que a manera de válvulas permiten el paso del l.c.r. hacia la luz del seno debido a la mayor presión de líquido en este espacio subaracnoideo.
Presión del l.c.r. cuando el individuo está en posición horizontal, la presión es la misma en todos los espacios subracnoideos craneovertebrales, con cifras entre 10 y 15 cm de agua y un poco menores dentro de los ventrículos en posición erecta o sentada la presión aumenta en el canal vertebral por el peso de la columna líquida hasta unos 30-50 cm de agua y se hace negativa en los ventrículos (-3 cm. de agua).
En la clínica la medida de la presión de l.c.r. se hace colocando un manómetro a un trocar de punción raquídea.
MENINGITIS
Inflamación de las meninges o membranas que envuelven el encéfalo y la médula espinal. La meningitis se clasifica como paquimeningitis cuando afecta a la duramadre (la membrana más externa), y como leptomeningitis cuando las membranas implicadas son la piamadre y la aracnoides (las más internas). Por lo general, la paquimeningitis es consecuencia de un traumatismo, como la fractura de los huesos del cráneo, o por la extensión de una infección localizada en el oído medio, la apófisis mastoides, los senos etmoidales o los senos frontales. La leptomeningitis es mucho más frecuente y se debe a: la propagación de una inflamación localizada en la nasofaringe; a la invasión de las meninges por microorganismos bacterianos a través de la circulación, como aquellos que producen la neumonía (neumococos); a un huésped de otros organismos, como el meningococo y Haemophilus influenzae.
La meningitis cerebroespinal epidémica o meningitis meningocócica es una enfermedad específica, muy infecciosa, que afecta al sistema respiratorio superior y a las meninges, en especial a niños y adultos jóvenes, y en particular en aquellos lugares donde convive mucha gente, como los dormitorios escolares o los cuarteles. Tiene una distribución casi mundial, aunque es más frecuente en las regiones templadas del norte. Las epidemias se han sucedido de forma periódica. El agente responsable es el meningococo, Neisseria meningitis, aislado en 1887 por el médico austriaco Anton Weichselbaum.
La mayoría de los casos de meningitis, en especial los producidos por bacterias, presentan un inicio brusco, con síntomas que incluyen cefalea, rigidez de nuca, fiebre, náuseas, vómitos, apatía e irritabilidad, que con frecuencia conducen a estupor y coma. Progresa con rapidez y si no se trata en periodos que oscilan entre 24 y 72 horas puede conducir a la muerte. La meningitis bacteriana se trata con mucha eficacia con la administración precoz de antibióticos.
A veces, en particular en los niños que no se tratan precozmente, se pueden producir lesiones cerebrales graves. Esto puede ocurrir cuando el drenaje del líquido cefalorraquídeo desde las cavidades hacia el cerebro se bloquea, y provoca distensión de las cavidades, presión sobre el cerebro, y aumento del cráneo, lo que se denomina HIDROCEFALIA.
La meningitis tuberculosa, que es más frecuente en niños que en adultos, suele ser secundaria a una tuberculosis pulmonar. Esta forma de meningitis fue casi siempre mortal hasta el descubrimiento de la estreptomicina. Desde la introducción de ésta, y de otros fármacos antituberculosos, se han producido muchas recuperaciones y las curaciones han sido eficaces incluso en casos recidivantes.
La meningitis viral generalmente es una forma no mortal de meningitis infecciosa. Casi siempre afecta a niños y origina síntomas como cefaleas, fiebre elevada, vómitos y dolor en las piernas. En la mayoría de las meningitis los pacientes afectados se recuperan de manera espontánea en un periodo de una a dos semanas.
HIDROCEFALIA.
Es una obstrucción de la circulación o absorción del l.c.r., o una producción mayor que el absorvido, que produce un aumento en la cantidad y la presión del l.c.r. lo cual es característico de la hidrocefalia.
El aumento del líquido en las cavidades ventriculares se donomina como hidrocefalia interna y puede ser debida a la obstrucción en el agujero interventricular, el acueducto o en los orificios del cuarto ventrículo. La acumulación del liquido en el espacio subracnoideo se donomina hidrocefalia externa.
25. IRRIGACION CEREBRAL
IRRIGACIÓN DEL CEREBRO
El cerebro está irrigado por las dos arterias carótidas internas y las dos arterias vertebrales. Las cuatro arterias se ubican dentro del espacio subaracnoideo y sus ramas se anastomosan en la superficie inferior del encéfalo formando el polígono de Willis.
Arteria carótida interna
La arteria carótida interna comienza en la bifurcación de la arteria carótida común (primitiva), donde habitualmente hay una dilatación localizada, denominada seno carotídeo. Asciende por el cuello y atraviesa la base del cráneo pasando a través del conducto carotídeo del hueso temporal. Luego la arteria discurre horizontalmente hacia adelante a través del seno cavernoso y sale en la cara medial de la apófisis clinoides anterior perforando la duramadre. Luego entra en el espacio subaracnoideo atravesando la aracnoides y gira hacia atrás hacia la región de la sustancia perforada anterior del encéfalo en el extremo interno de la cisura lateral. Aquí se divide en las arterias cerebrales anterior y media.
Ramas de la porción cerebral de la arteria carótida interna
1. La arteria oftálmica se origina donde la arteria carótida interna sale del seno cavernoso. Entra en la órbita a través del agujero óptico por debajo y por fuera del nervio óptico. Irriga el ojo y otras estructuras orbitarias y sus ramas terminales irrigan el área frontal del cuero cabelludo, los senos etmoidal y frontal y el dorso de la nariz.
La arteria comunicante posterior es un pequeño vaso que se origina en la arteria carótida interna cerca de su bifurcación terminal. La arteria comunicante posterior discurre hacia atrás por encima del nervio oculomotor para unirse con la arteria cerebral posterior, formando parte del polígono de Willis.
La arteria coroidea anterior, una rama pequeña, también se origina en la arteria carótida interna cerca de su bifurcación terminal. Se dirige hacia atrás cerca de la cintilla óptica, entra en el asta inferior del ventrículo lateral y termina en el plexo coroideo. Da origen a numerosas ramas pequeñas para las estructuras circundantes, que incluyen el pie del pedúnculo cerebral, cuerpo geniculado lateral, cintilla óptica y también cápsula interna.
La arteria cerebral anterior es la rama terminal más pequeña de la arteria carótida interna. Se dirige hacia delante medialmente por encima del nervio óptico y entra en la cisura longitudinal del cerebro. Aquí se une con la arteria cerebral anterior del lado opuesto por medio de la arteria comunicante anterior. Se curva hacia atrás sobre el cuerpo calloso y finalmente se anastomosa con la arteria cerebral posterior.
Las ramas corticales irrigan toda la superficie medial de la corteza cerebral hasta la cisura parietooccipital. También irrigan una tira de corteza de aproximadamente 2,5 cm de ancho en la superficie lateral adyacente. Así, la arteria cerebral anterior irriga el "área de la pierna" de la circunvolución precentral. Un grupo de ramas centrales atraviesa la sustancia perforada anterior y ayuda a irrigar p artes de los núcleos lenticular y caudado la cápsula interna.
5. La arteria cerebral media, la rama más grande de la carótida interna, discurre lateralmente en la cisura lateral. Ramas corticales irrigan toda la superficie lateral del hemisferio, excepto la estrecha banda irrigada por la arteria cerebral anterior, el polo occipital y la cara Arteria vertebral inferolateral del hemisferio que están irrigados por la arteria cerebral posterior. Así, esta arteria irriga toda el área motora excepto el "área de la pierna . Ramas centrales entran en la sustancia perforada anterior e irrigan los núcleos lentiforme y caudado y la cápsula interna. La arteria vertebral, rama de la primera parte de la arteria subclavia, asciende por el cuello atravesando los agujeros en las apófisis transversas de las seis vértebras cervicales superiores. Entra en el cráneo a través del agujero occipital y atraviesa la duramadre y aracnoides para entrar en el espacio subaracnoideo. Luego transcurre hacia arriba, hacia adelante y en dirección medial en el bulbo raquídeo. En el borde inferior de la protuberancia, se une con el vaso del lado opuesto para formar la arteria basilar.
Ramas de la porción craneal de la arteria vertebral
1. Las ramas meníngeas son pequeñas e irrigan el hueso y la duramadre en la fosa craneal posterior.
2. La arteria espinal posterior puede originarse en la arteria vertebral o en la arteria cerebelosa posteroinferior. Desciende sobre la cara posterior de la médula espinal cerca de las raíces posteriores de los nervios espinales. Las ramas están reforzadas por las arterias radiculares que entran en el conducto raquídeo a través de los agujeros intervertebrales.
3. La arteria espinal anterior se forma a partir de una rama contribuyente de cada arteria vertebral cerca de su terminación. La arteria única desciende en la cara anterior del bulbo raquídeo y la médula espinal y está incluida en la piamadre a lo largo del surco medio anterior. La arteria es reforzada por arterias radiculares que entran en el conducto raquídeo a través de los agujeros intervertebrales.
4. La arteria cerebelosa posteroinferior, la rama más grande de la arteria vertebral, sigue un curso irregular entre el bulbo raquídeo y el cerebelo. Irriga la cara inferior del vermis, los núcleos centrales del cerebelo y la cara inferior del hemisferio cerebeloso; también irriga el bulbo raquídeo y el plexo coroideo del cuarto ventrículo.
5. Las arterias bulbares son ramas muy pequeñas que se distribuyen en el bulbo raquídeo.
Arteria basilar
La arteria basilar, formada por la unión de las dos arterias vertebrales, asciende en un surco en la cara anterior de la protuberancia. En el límite superior de la protuberancia se divide en las dos arterias cerebrales posteriores.
Ramas de la arteria basilar
1. Las arterias protuberanciales son numerosos vasos pequeños que entran en la sustancia de la protuberancia.
2. La arteria laberíntica (auditiva interna)
Es una larga arteria estrecha que acompaña a los nervios facial y vestibulococlear en el conducto auditivo interno e irriga el oído interno. A menudo nace de la arteria cerebelosa anteroinferior.
3. La arteria cerebelosa anteroinferior se dirige hacia atrás y lateralmente e irriga las partes anterior e inferior del cerebelo. Unas pocas ramas se dirigen a la protuberancia y la parte superior del bulbo raquídeo.
4. La arteria cerebelosa superior se origina cerca de la terminación de la arteria basilar. Pasa alrededor del pedúnculo cerebral e irriga la superficie superior del cerebelo. También irriga la protuberancia, el cuerpo pineal y el velo medular superior.
5. La arteria cerebral posterior se curva lateralmente y hacia atrás alrededor del mesencéfalo y se une con la rama comunicante posterior de la arteria carótida interna. Ramas corticales irrigan las superficies inferolateral y medial del lóbulo temporal y las superficies lateral y medial del lóbulo occipital. Así, la arteria cerebral posterior irriga la corteza visual. Ramas centrales penetran en la sustancia cerebral e irrigan partes del tálamo y del núcleo lentiforme y el mesencéfalo, cuerpo pineal y cuerpo geniculado interno. Una rama coroidea entra en el asta inferior del ventrículo lateral e irriga el plexo coroideo; también irriga el plexo coroideo del tercer ventrículo.
El polígono de Willis se ubica en la fosa interpeduncular en la base del encéfalo. Está formado por la anastomosis entre las dos arterias carótidas internas y las dos arterias vertebrales. Las arterias comunicantes anteriores, cerebrales anteriores, carótidas internas, comunicantes posteriores, cerebrales posteriores y basilar contribuyen a formar el polígono. El polígono de Willis permite que la sangre que entra por cualquiera de las arterias carótidas internas o vertebrales sea distribuida hacia cualquier parte de ambos hemisferios cerebrales. Ramas corticales y centrales se originan en el polígono e irrigan la sustancia encefálica.
Arterias hacia áreas encefálicas específicas
El cuerpo estriado y la cápsula interna están irrigados principalmente por las ramas estriadas internas y externas de la arteria cerebral media; las ramas centrales de la arteria cerebral anterior irrigan el resto de estas estructuras.
El tálamo está irrigado principalmente por ramas de las arterias comunicante posterior, basilar y cerebral posterior.
El mesencéfalo está irrigado por las arterias cerebrales posteriores, cerebelosas superiores y basilar.
La protuberancia está irrigada por las arterias basilar y cerebelosas anteroinferiores y superiores.
El bulbo raquídeo está irrigado por las arterias vertebrales, espinales anterior y posterior, cerebelosas posteroinferiores y la arteria basilar.
El cerebelo está irrigado por las arterias cerebelosas superiores, anteroinferiores y posteroinferiores.
Inervación de las arterias cerebrales
Las arterias cerebrales reciben una rica inervación de fibras nerviosas simpáticas posganglionares. Estas fibras se originan en el ganglio simpático cervical superior. La estimulación de estos nervios produce vasoconstricción de las arterias cerebrales. Sin embargo, en condiciones normales, el flujo sanguíneo local es controlado principalmente por las concentraciones de dióxido de carbono, hidrogeniones y oxígeno presentes en el tejido nervioso; una elevación en las concentraciones de dióxido de carbono e hidrogeniones y una reducción de la tensión de oxigeno producen una vasodilatación.
VENAS DEL ENCÉFALO
Las venas del encéfalo no tienen tejido muscular en su paredes muy delgadas ni poseen válvulas. Salen del encéfalo y se ubican en el espacio subaracnoideo. Atraviesan la aracnoides y la capa meníngea de la duramadre y drenan en los senos venosos craneales.
Venas cerebrales externas
Las venas cerebrales superiores se dirigen hacia arriba sobre la superficie lateral del hemisferio cerebral y drenan en el seno sagital superior (fig. 28-5).
La vena cerebral media superficial drena la superficie lateral del hemisferio cerebral. Discurre por debajo de la cisura lateral y se vacía en el seno cavernoso (fig. 28-5).
La vena cerebral media profunda drena la ínsula y se une con las venas cerebral anterior y del cuerpo estriado para formar la vena basal. La vena basal finalmente se une con la vena cerebral magna, la cual drena en el seno recto (fig. 28-5).
Venas cerebrales internas
Hay dos venas cerebrales internas y están formadas por la unión de la vena talamoestriada y la vena coroidea en el agujero interventricular. Las dos venas discurren posteriormente en la tela coroidea del tercer ventrículo y se unen por debajo del rodete del cuerpo calloso para formar la vena cerebral magna, la cual drena en el seno recto.
Venas de áreas encefálicas específicas
El mesencéfalo es drenado por venas que desembocan en las venas básales o cerebrales magnas.
La protuberancia es drenada por venas que desembocan en las venas básales, venas cerebelosas o senos venosos vecinos.
El bulbo raquídeo es drenado por venas que desembocan en las venas espinales y senos venosos vecinos.
El cerebelo es drenado por venas que desembocan en las venas cerebrales magnas o senos venosos adyacentes.
Descargar
| Enviado por: | El remitente no desea revelar su nombre |
| Idioma: | castellano |
| País: | Colombia |
Todos los derechos reservados.