Biología, Botánica, Genética y Zoología
Equinodermos
INTRODUCCION
Los equinodermos constituyen un grupo zoológico marino caracterizado por una simetría radial de orden 5 en el estado adulto (simetría penta radial), esta simetría, se adquiere tras una metamorfosis que se produce al final del desarrollo embrionario y larval.
Los equinodermos actuales se dividen en cuatro clases, echinoideos (erizos de mar, aún llamados: "castañas de mar"), los asteroideos (estrellas de mar), los holoturoideos (pepinos de mar) y los ofiuroideos. Ahora bien, por qué el interés mayor en los echinoideos? Bien, si comenzamos a describirlo, diremos que tiene un esqueleto formado por placas soldadas, y un aparato masticador ubicado en el polo oral llamado la “linterna de Aristóteles” posee también muchos podios que salen de unos orificios microscópicos los cuales le sirve ara trasladarse. El interés para el estudio de erizos de mar reside en el hecho de que los ovarios de estos animales pueden producir decenas de millares de ovocitos, su diámetro no es tan pequeño, lo que los vuelve accesible al experimentador bajo un simple microscopio. Por otro lado, la abundancia de algunas especies sobre nuestras costas hace su suministro fácil. Desde el punto de vista embriológico, las fases del desarrollo son fáciles de observar. Las etapas sucesivas claramente separadas y cuyos mecanismos de desarrollo son al parecer simples de comprender hacen de este un organismo sencillo. Por último, la cronología del desarrollo precoz hasta la organogénesis no tarda más de tres días a la temperatura de 18°C.
| estadios | tiempo para casa uno de los estadios |
| 2 células | 1h30-2h |
| 4 células | 2h30-3h |
| 4h30 | |
| 64 células | 5h30 |
| 128 células | 6h30-7h |
| blástula | 14h |
| gastrula) | 24h |
| larva prisma | 30h-35h |
| larva pluteus joven | 50h-55h |
CAPITULO I
Los gametos
En los erizos de mar, los sexos son separados. Los productos genitales Son expulsados al exterior, y la fecundación es fuera, el producto genital en las hembras es anaranjada y blanca en los varones.
|
|

Los productos genitales proceden de las gónadas. Obedeciendo a la simetría pentaradial de estos animales, los gónadas son 5 y dispuestos de manera irradiada con relación al eje polo oral- polo aboral.
|
|
| Disección de la región aboral |

El ovocito
El número de ovocitos puestos por una hembra puede ser del orden de la decena de millones y representa un volumen de 10 a 20 mililitros.
En el ovario, el ovocito posee un núcleo de gran tamaño llamado vesícula germinativa en la que se puede distinguir un nucleolo.
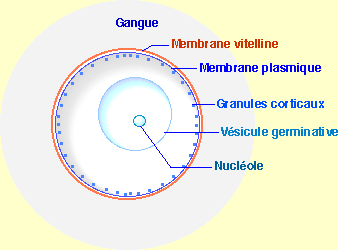
Esquema de interpretación del óvulo
En el erizo de mar, el ovocito presenta una pigmentación anaranjada uniforme. Por otra parte, los erizos de mar hembras a madurez se reconocen por la emisión de sus productos genitales anaranjados. El pigmento se reunirá en forma de un anillo en el hemisferio vegetativo después de la fecundación. Es la señal exterior de la polaridad axial polo animal- polo vegetativo. Se habla entonces de hemisferio animal no pigmentado y de hemisferio vegetativo pigmentado.
|
|
| La celula no fecundada posee una coloracion uniforme, mientras que la fecundada, es como se ve en a figura |

El espermatozoide
El espermatozoide es una célula móvil, de pequeño tamaño (algunos micrones), con un citoplasma muy reducido. Incluye tres partes principales:
- la cabeza con el núcleo,
- la parte intermedia con los centríolos y el conjunto de las mitocondrias
- la cola o flagelo, órgano de motricidad.
|
|
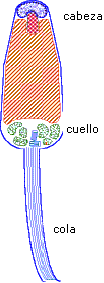
La fecundación
En una región dada, todos los individuos de una misma población de erizo de mar se reproducen en el mismo momento. Los productos genitales pues son emitidos por millones en el agua de mar. Estas poblaciones de erizo de mar que es generalmente muy numerosas, son, en realidad, miles de millones de ovocitos y espermatozoides que se liberan. Los movimientos del agua de mar y las corrientes contribuyen a la mezcla de los gametos y favorecen su encuentro. Además, mecanismos moleculares específicos de atracción de los espermatozoides (quimiotactismo) y de interacción celular (adhesión del espermatozoide) preparan al espermatozoide a su reconocimiento por el ovocito. La especificidad de estas interacciones contribuye por otra parte a limitar los riesgos de fecundación cruzada interespecífica.
Tales concentraciones de gametos pueden implicar otro riesgo, aquél que un mismo ovocito esté fertilizado por varios espermatozoides. Esto es lo que se llama polispermia. Ahora bien, veremos que mecanismos subcelulares se oponen a esta anomalía. Son las etapas de los bloqueos de la polispermia
.
En total, la fecundación desencadena una cascada de acontecimientos de la que cada etapa depende de la anterior. Esta secuencia, como por otra parte más generalmente el desarrollo embrionario, es la traducción de la aplicación de un programa genético del desarrollo.
La fecundación aparece como una cascada de acontecimientos celulares que se refieren en primer lugar al espermatozoide luego el ovocito, prensando ahora las 5 etapas:
1.- Atracción del espermatozoide
2.- Reacción acrosómica
3.- Adhesión del espermatozoide
4.- Activación del huevo (bloqueo precoz de la polispermia)
5.- Reacción cortical (bloqueo tardío de la polispermia)
La division celular
Caracteriza el paso del estado unicelular al estado pluricelular dado por mitosis sucesivas y rápidas que conducen a la formación de la blástula. Cada blastomero sufre una bipartición en células jóvenes. La segmentación es total, igual y meridional hasta la fase 8 blastómeros, luego se vuelve desigual a partir de la fase 16 blastómeros. Al término de la segmentación, la blástula se compone de cerca de 2000 blastomères. Progresivamente, una cavidad central se forma, la cavidad de segmentación o blastócele.
2 blastómeros
El primer surco de separación aparece alrededor de una hora treinta después de la fecundación, a la temperatura de 15°C. Una constricción meridional resulta al mismo tiempo que nuevas membranas plásmicas son elaboradas por los dos primeros blastómeros que nacen.
4-8 blastómeros
El segundo surco de separación es también meridiano y perpendicular al primero, separando cuatro células de dimensiones idénticas. El tercer surco de separación es ecuatorial y perpendicular a los dos precedentes. Se forman ocho células similares.
16 blastomères
A partir de la fase 16 blastómeros, las divisiones celulares se convierten en desiguales. En el hemisferio animal, los planes de separación son meridianos y generan ocho células similares. En el hemisferio vegetativo, los planes de separación son latitudinales y claramente compensados hacia el polo vegetativo. Dos clases de células muchachas son resultantes: los macrómeros próximos a Ecuador y los micromèros al polo
Vegetativo.
La diferencia de importancia entre los macrómeros y los micromèros resulta de una mitosis asimétrica. La disimetría macrómeros/micromèros está vinculada a una disimetría en el eje mitótico. Se puso de manifiesto que los centrosomas de los ejes mitoticos vegetativos son mismos ellos de tamaño desigual. Así el centrosoma situado del lado del polo vegetativo es claramente más pequeño que del centrosoma situado hacia el Ecuador del embrión.

| Disimetría de orientación |
Blástula
A partir de la fase 32 blastómeros, la segmentación desigual se esfuma. Es en esta fase que aparece una cavidad de segmentación central, el blastocele. A la fase 128 células, la blastula es formado por un epitelio unistratificado y esférico. Es limitado por una membrana básica interna y la capa hialina externa. Uniones apretadas contribuyen a mantener la cohesión entre las células.
|
|
| Joven blastula. |
La cara interna de las células se empapela de una estructura extracelular formada por glicoproteínas de alto peso molecular, la matriz extracelular.
Al final de la segmentación, el blástula ciliada enteramente. El polo animal lleva un penacho de latigazos más largos difíciles a poner de relieve.
El huevo de erizo de mar es cualitativamente heterogéneo. En morfología externa, ya se sabe que la distribución del pigmento cortical anaranjado así como la diferencia de importancia entre los blastómeros de la fase 16 células son una indicación de esta heteregeneidad. Al final de la segmentación, la blástula se forma por un mosaico de cinco territorios presuntos dispuestos de manera concéntrica en torno al eje polo animal- polo vegetativo. En el orden polo animal- polo vegetativo, se distinguen los siguientes territorios:
- Animal1 (An1), en el origen de la epidermis de los brazos orales.
- Animal2 (An2), en el origen de la epidermis de los brazos anales.
- Vegetativo1 (Vg1), en el origen de la epidermis de los brazos anales y de la cara anal así como una parte del endodermo.
- Vegetativo2 (Vg2), en el origen del endodermo y mesenquima del secundario.
- Micromeros, en el origen del mesenquina enseñanza primaria.
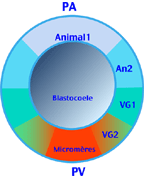
| Carta de territorios |
El embrión posee movimiento y se desplaza en el plancton gracias a sus cilios.
En resumen, al término de la segmentación, el embrión esférico presenta un epitelio unistratificado en torno a una amplia cavidad de segmentación, el blastocele. Una matriz extracelular que empapela la pared el blastocele y la capa hialina. El mapa de los territorios presuntos nos indica como se determina número de éstos, en particular, los micromèros al polo vegetativo. Por lo tanto, es a partir de esta región que la gastrulación se iniciará y los micromèros serán las primeras células por animarse.
Blástula al término de su estadio
El gastrulacion
El gastrulation es caracterizado por:
1.- Aparición de los movimientos celulares coordinados. (movimientos
morfogenéticos).
2.- La formación de los tres tejidos fundamentales: el ectodermo, dando origen de la epidermis, el mesodermo dando origen a los mésenquimas primario y secundario, y el endodermo dando origen del tubo digestivo.
3.- Aparición de una cavidad secundaria digestiva, el arquenterón.
En paralelo, el ritmo amitótico disminuye y el ciclo celular se alarga.
Análisis de los movimientos celulares durante el gastrulación
Se pueden clasificar los movimientos celulares aplicados durante la gastrulacion del erizo de mar en 4 categorías. Cada uno de estos movimientos está vinculado a una etapa de la gastrulacion:
a- Ingreso del mesenquima.
b- Migración del mésenquima.
c- Inflexión del polo vegetativo en del blastocèle, quién inicia la formación del arquentéron.
d- Alargamiento del arquentéron e individualización mesenquima
a - Ingreso del mesenquima
|
|
|
|
| Esquema de interpretación de ingreso del mesenquima |
| Esquema de ingreso del mesenquima primario |
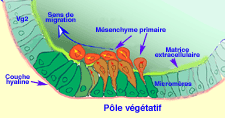

b - Migracion del mésenquime primario
Las células en migración entran en contacto con la matriz extracelular, sobre la pared del blastocoele. Varias moléculas desempeñan un papel importante en estos acontecimientos.

migración de celulas del mesenquima primario
c - Inflexión del polo vegetativo en del blastocoele
|
|
|
|
| Momento de inflexion | corvatura | Esquema del momento de inflexion en polo vegetativo |



La inflexión del polo vegetativo constituye el principio del invaginación del endodermo, y el panorama del arquentéron abierto sobre el exterior por del blastoporo. No sobrepasa 1/4 a 1/3 de la distancia polo animal- polo vegetativo. Esto quiere decir que este movimiento no es suficiente para garantizar la formación completa del arquentéron y no permite, en particular, su extensión hacia el polo animal.
d - Alargamiento del archentéron e individualización mésenquima
|
|
|
|



| Elongación el arquenteron e individualización del mesenquima |
El alargamiento del arquenterón está garantizado por la llanura de las células que componen sus paredes. Además, movimientos de intercalación celular reducen el diámetro del arquenterón para hacer un estrecho tubo y alargado. Se habla de movimientos de convergencia extensión especialmente netos en la estrella de mar
Otro mecanismo puede contribuir a este alargamiento. Las células situadas al ápice del arquenterón emiten filopodes, a veces muy largos, puesto que van al encuentro del techo del balstocoele.
Una vez esta conexión garantizada, los fipodes se contratan, ejerciendo una fuerza de tracción que contribuye a la prolongación del endodermo hacia el polo animal. Estos movimientos se refieren siempre al vegetativo.
CICLO LARVAL
A salir del gastrulation, no se terminan los movimientos morfogenéticos. El modelado de los tejidos de la gastrula a conducido al embrión hacia la fase larval. En primer lugar, durante su alargamiento, la extremidad apical del arquentéron se desciende hacia la futura cara oral. Esta región parece determinarse muy pronto durante el desarrollo.
Las células del mesenquima secundario tienen un papel determinante en el proceso de inflexión. En efecto, los largos filipodes de las células del mesenquima secundario que participan en el alargamiento del arquenterón tendrían también por papel reconocer la cara ventral de la epidermis durante su exploración de la pared del blastocele. Una vez esta información recogida, las células del mesénquima secundario conducirían la extremidad previa del arquenterón al contacto de la epidermis ventral en el origen de la región oral de la larva. La boca se abre al contacto del ápice del arquenterón con la epidermis.
Sabiendo que el blastoporo está destinado a convertirse en el ano, la apertura secundaria de la boca califica a los equinodermos de deurostomados a los cuales pertenecen también cordados. Continuando, a región oral se aplana. Se le da el nombre de larva prisma.
La boca se centra en una depresión dicha dipleurula, los que bordes se surten de una tira ciliada. La larva es un dipleurula. Esta depresión aumenta y la tira ciliada forma extensiones apoyadas por las espículas en crecimiento.
De estas extensiones nacen cuatro "brazos" cuyos dos crecen del lado de la boca y dos del lado del ano (los brazos orales y los brazos anales). Posteriormente, la reunión de las espículas forma un haz que da una forma alargada a la larva que se llama ahora larva pluteus,
En el interior de la larva, el arquenterón se divide en tres regiones. En el sentido antero posterior, se distingue un corto esófago que desemboca en un bolsillo hinchado, el estómago, al cual hace consecuencia el tubo intestinal abierto en el ano.
La larva plutéus es nadadora y planctónica. Avanza dirigiendo su cara oral hacia el frente. El pluteus se alimenta con diatomeas planctónicas.

La metamorfosis
En el erizo de mar, en el momento en que el archentéron entra en contacto con la epidermis para formar la boca, en su extremidad previa florece una vesícula celómica que se divide en bolsos celómicas derechos e izquierdos. Éstas se colocan de manera simétrica de cada lado del tubo digestivo.
| Detalle del pluteus en vista lateral | Vista ventral |
Cada vesícula celómica sufre un metamerisacion tripartito; previa, intermediario y posterior. Las vesículas celómicas posteriores rodean el bolsillo estomacal para formar el conducto gástrico. La vesícula celómicas intermedia izquierda toma el nombre de hidrocele (en el origen del sistema acuífero de adulto), mientras que el equivalente derecho sufre una fuerte reducción. La vesícula celomicae previa izquierda florece un canal que se dirige hacia la epidermis y desemboca en el medio exterior. Se trata del canal madrepórico.
En paralelo, una conexión se establece entre el conducto previo izquierdo y el hidrocele por la formación de un canal que pone en comunicación las dos vesículas. Se trata del futuro canal de la arena. Por último, la epidermis al contacto con el hidrocele se invagina y forma un bolsillo destinado a englobar hidrocele. Por analogía con los mamíferos, se da a esta estructura el nombre de amnios. Es a partir de este conjunto que se efectuará la metamorfosis del erizo de mar.
El bolsillo amniótico así formado con los derivados celómicos toma el nombre de bolso. Dentro, el hidrocele evoluciona en anillo y florecen cinco divertículos. El conjunto se vuelve pentagonal y es, a partir de esta estructura, que el joven erizo de mar se construye. Aparte del bolso, el estómago, los celomas previos y posteriores izquierdas así como el ectodermo dorsal del pluteus participan en esta metamorfosis.
En consecuencia, mientras que la metamorfosis se continúa, el pluteus lleva en él a un joven erizo de mar miniatura surtido de algunas espinas y de cinco podias primarias. Luego la cavidad amniótica se abre, el erizo de mar introduce el resto del pluteus no utilizado para la metamorfosis del joven. Por último, a medida que el conjunto se sobrecargado por el esqueleto calcáreo en formación así como por las primeras espinas, el erizo de mar desciende hacia el fondo marino.
Índice
Introducción
Objetivos
Capitulo I
Los gametos
Ovocito y Espermatozoide
Fecundación
División celular
Ciclo larval
Metamorfosis
Capitulo II
Materiales y metodos
Resultados
Recomendaciones
Conclusiones
Bibliografía
Descargar
| Enviado por: | Trilce |
| Idioma: | castellano |
| País: | Perú |
Todos los derechos reservados.