Salud
Bases biofísicas del electrocardiograma
SEMINARIO DE BIOFISICA
TEMA: BASES BIOFISICAS DEL ELECTROCARDIOGRAMA
PERU
2006
INTRODUCCION
OBJETIVOS
GENERAL
Saber interpretar un electrocardiograma normal.
MARCO TEORICO
BASES BIOFISICAS DEL ELECTROCARDIOGRAMA
1.-ANATOMOFISIOLOGIA DEL MÚSCULO
Fibra del músculo esquelético
Están formados por numerosas fibras cuyos diámetros varían entre 10 y 80 micrómetros. Cada una de estas fibras está formada a su vez por subunidades sucesivamente más pequeña, también mostradas.
Cada una está inervada pro una sola terminación nerviosa, localizada cerca de su porción media.
SARCOLEMA. El sarcolema es la membrana celular de la fibra muscular. Está constituido por una membrana celular verdadera, denominada membrana plasmática, y por una cubierta exterior formada por una fina capa de material polisacárido que contiene numerosas fibrillas finas de colágeno. En cada extremo de la fibra muscular, esta capa superficial del sarcolema se fusiona con una fibra tendinosa, y estas fibras tendinosas se unen a su vez en hacer para formar los tendones musculares e insertarse en los huesos.
MIOFIBRILLAS; FILAMENTOS DE ACTINA Y DE MIOSINA. Cada fibra muscular contiene ente varios centenares y varios millones de miofibrillas. Cada miofibrilla posee a su vez, situados uno al lado de otro, unos 1500 filamentos de misina y 3000 filamentos de actino, que son grandes moléculas proteínas polimerizadas responsables de la contracción muscular.
Los filamentos de actina y de miosina están parcialmente intercalados, lo que hace que las miofibrillas presenten bandas clareas y oscuras alternantes, las bandas claras contienen tan solo filamentos de actina y se denominan bandas I porque son isotrópicas con la luz polarizada. Las bandas oscuras contienen los filamentos de miosina, así como los extremos de los filamentos de actina donde estos se superponen a la misina, y se denominan bandas A porque son anisotrópicas con la luz polarizada. Obsérvense también estas estructuras son los puentes cruzados, y hacen prominencia desde las superficies de los filamentos de miosina a lo largo de toda la extensión del filamento, excepto en el centro exacto de éste. La interacción entre estos puentes cruzados y los filamentos de actina produce la contracción.
Los filamentos de actina están unidos a un denominado disco Z. desde este disco, estos filamentos de miosina. El disco y misosina, cruza la miofibrilla y también cruza de una miofibrilla a otra, uniéndolas ente si a todo lo ancho de la fibra muscular.
La porción de una mifibrilla (o detona la fibra muscular) situada entre dos discos Z sucesivos se denomina sarcómero.
¿Qué MANTIENE LOS FILAMENTOS DE MIOSINA Y ACTINA EN POSICÓN ? MOLÉCULAS FILAMENTOSAS DE TITINA.
La relación lado a lado ente los filamentos de actina y misina es difícil de mantener. Esto se logra mediante un gran número de moléculas filamentosas de una proteína denominada titina.
Además, debido a que es filamentosas, es una molécula muy elástica. Estas elásticas moléculas de titina actúan como armazón que reviste los filamentos de actina y miosina para constituir la maquinaria contráctil de trabajo del sarcómero.
SARCOPLASMA. Dentro de la fibra muscular, las miofibrillas están suspendidas en una matriz denominada sarcoplasma, constituida por los componentes intracelulares habituales. El líquido del sarcoplasma contiene grandes cantidades de potasio, magnesio y fosfato, además de numerosas enzimas proteicas. También está presente un elevado número de mitocondrias, situdas paralelas a las miofibrillas situación indicada de la gran necesidad de las mifibrillas contráctiles de disponer.
RETÍCULO SARCOPLÁSMICO. También en el sarcoplasma existe un extenso reticulo endoplásmico que en la fibra muscular se denomina retículo sarcoplásmico.
Los tipos de músculo de contracción más rápida poseen retículos sarcoplásmicos especialmente extensos, lo que indica.
MECANISMO GENERAL DE LA CONTRACCIÓN MUSCULAR
La iniciación y la ejecución de la contracción muscular se producen según los siguientes pasos secuenciales.
Un potencial de acción viaja a lo largo de un nervio motor hasta sus terminaciones en las fibras musculares.
En cada terminación, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
la acetilcolina actúa sobre una zona local de la membrana de la fibra muscular para abrir múltiples canales con apertura por acetilcolina a través de moléculas proteicas que flotan en la membrana.
la apertura de los canales de cetilcolina permite que grandes cantidades de iones sodio fluyan al interior de la membrana de la fibra muscular. Esto inicia un potencial de acción en la fibra muscular.
El potencial de acción viaja a lo largo de la membrana de la fibra muscular.
el potencial de acción despolariza la membrana de la fibra muscular y gran parte de la electricidad del potencial de acción también viaja en profanidad dentro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que estaban almacenados en el retículo.
los iones calcio inician fuerzas de atracción entre los filamentos de actina y de miosina, haciendo que se deslicen entre sí, lo cual constituye el proceso de contracción.
transcurrida una fracción de segundo, los iones calcio son bombeados de nuevo al interior del retículo sarcoplásmico mediante una bomba de calcio de membrana.
MECANISMO MOLECULAR DE LA CONTRACCIÓN MUSCULAR
La contracción muscular se produce por un mecanismo de deslizamiento de los filamentos.
Características moleculares de los filamentos contráctiles
Filamentos de miosina.
La molécula de miosina está compuesta por seis cadenas polipertídicas, dos cadenas pesadas, cada una con un peso molecular de unos 200.000, y cuatro cadenas ligeras, con pesos moleculares de alrededor de 20.000 cada una las dos cadenas pesadas se enrollan en espiral entre sí para formar una sobre hélice, denominada de la molécula de miosina. Un extremo de cada una de estas cadenas está plegado en una estructura polipeptídica globulosa denominada cabezas de la miosina.
El filamento de miosina está constituido por 200 o más moléculas individuales de miosina. Así mismo, parte de la porción de la hélice de cada molécula de miosina se extiende hacia el lateral junto con la cabeza, proporcionando así un brazo que aparta la cabeza del cuerpo hacia el exterior, salientes se denominan en conjunto puntes cruzados.
La longitud total de cada filamento de miosina es uniforme, casi exactamente 1.6 micrómetros
FILAMENTO DE ACTINA. El filamento de actina también es complejo. Está constituido por tres componentes proteicos: actina, tropomiosina y troponina.
La columna vertebral del filamento de actina es una molécula proteica de actina F de doble hebra.
Cada filamento de la doble hélice de actina F está compuesto por moléculas de actina G polimerizadas.
Cada filamento de actina tiene alrededor de 1 micrómetro de logitud. Las bases de estos filamentos están fuertemente insertadas en dos discos Z, mientras que los extremos sobresalen en ambas direcciones para situarse en los espacios existentes entre las moléculas de miosina.
MOLÉCULAS DE TROPOMIOSINA
Estas moléculas están conectadas enrolladas en espiral alrededor de los lados de la hélice de actina F. en estado de reposo, las moléculas de tropomiosina descansan sobre los sitios activos de las hebras de actina.
LA TROPONINA
Se trata en realidad de complejos de tres subunidades proteicas unidas de forma laxa, cada una de las cuales tiene un papel especifico en el control de la contracción muscular Una de las subunidades (troponina) posee gran afinidad por la actina, otra (troponina T) por la tropomiosina y la tercera (troponina C) por los iones calcio.
INTERACCIÓN DE LA MIOSINA, LOS FILAMENTOS DE ACTINA Y LOS IONES CALCIO PARA PRODUCIR LA CONTRACCIÓN.
Efectos de la desnervación muscular
Por tanto, la atrofia comienza casí inmediatamente. En la fase final de la atrofia por desnervación, la mayor parte de las fibras musculares está destruida y reemplazada por tejido fibroso y graso.
El tejido fibroso que reemplaza a las fibras musculares durante la atrofia por desnervación tiene tendencia a continuar acortándose durante muchos meses, lo que se denomina contractura.
RECUPERACIÓN DE LA CONTRATACIÓN MUSCULAR EN LA POLIOMIELITIS:
Poliomielitis, las fibras nerviosas restantes hacen brotar nuevos axones para formar muchas nuevas ramas que posteriormente enervan muchas de las fibras nerviosas paralizadas.
RIGOR MORTIS
Estado de contractura “rigor mortis”
MÚSCULO CARDIACO
MORFOLOGÍA
Las estriaciones del músculo cardiaco son similares alas del esquelético, y tiene líneas Z. Grandes cantidades de mitocondrias alargadas, están en estrecho contacto con las fibrillas musculares. Las fibras musculares se ramifican y se intercalan aunque cada una es una unidad completa, rodeada por una membrana celular. En el sitio donde el extremo de una fibra muscular colinda con otra, las membranas de ambas fibras se disponen en forma paralela entre si mediante una extensa serie de pliegues. Estas áreas, que siempre se encuentran en las líneas Z, se llaman discos intercalares estos brindan una unión firme entre las fibras, lo cual mantiene la cohesión entre las células.
El sistema T del músculo cardiaco se localiza en las líneas Z, en lugar de las uniones entre las bandas A e I, como sucede en el músculo esquelético de los mamíferos.
PROPIEDADES ELÉCTRICAS
Potenciales de membrana en reposo y de acción
El potencial de membrana en reposo de las células musculares cardiacas individuales de los mamíferos es cercano a 90 Mv interior es negativo con respecto al exterior de dos meses. Pero la fa se de meseta y repolarización dura 200 mseg o más. Como en otros tejidos excitables, los cambios en al concentración externa de K afectan el potencial de membrana en reposo del músculo cardiaco, en tanto que las modificaciones en la concentración externa de Na afectan la magnitud del potencial de acción. La despolarización rápida inicial y la inversión del potencial (fase O) se deben a la abertura de los canales del Na activados por voltaje, de manera similar a lo que ocurre en el nervio y en el músculo esquelético. La repolarización rápida inicial se produce por el cierre de los canales para el Na la siguiente meseta prolongada (fase 2) es resultado de una abertura más lenta, pero prolongada, de los canales de Ca2+ activados por voltaje. La repolarización final (fase 3) hasta el potencial de membrana en reposos (fase 4) se debe al cierre de los canales del Ca2+ con salida de K+ a través de varios tipos de canales para este ión.
PROPIEDADES MECÁNICAS
Respuesta contráctil
La respuesta contráctil del músculo cardiaco empieza justo después del inicio de la despolarización, y dura alrededor de 1.5 veces más que el potencial de acción. La función del Ca2+ en el acoplamiento de excitación con tracción es similar ala que tiene en el músculo esquelético.
HIPERTROFIA CARDIACA
Al igual que el músculo esquelético, el cardiaco sufre hipertrofia cuando hay aumento crónico en la carga, como sucede, por ejemplo, en la hipertensión. También existe hipertrofia en presencia de mutaciones en los genes que codifican para las diversas proteínas que constituyen el aparato contráctil, lo cual debilita la fuerza de contracción y produce miocardiopatía hipertrófica.
Efecto de los iones potasio y calcio sobre la función cardiaca
EFECTO DE LOS IONES POTASIO.
El exceso de potasio en los líquidos extracelulares hace que el corazón se dilate y quede flácido, y que también disminuya la frecuencia caradíaca. Las cantidades grandes pueden también bloquear la conducción del impulso cardíaco de las aurículas a los ventrículos a través del haz A-V.
Estos efectos son producidos en parte porque si existe una concentración elevada de potasio en los líquidos extracelulares, disminuye el potencial de membrana de reposo de las fibras musculares cardíacas.
EFECTO DE LOS IONES CALCIO.
El exceso de iones calcio produce efectos casí exactamente opuestos a los de los iones potasio, haciendo que el corazón caiga en una contracción espástica.
A la inversa, el déficit de iones calcio causa flacidez cardíaca, similar al efecto del potasio elevado. Afortunadamente, los niveles sanguíneos de calcio iónico, están normalmente regulados en una banda muy están normalmente regulados en una banda muy estrecha.
EFECTO DE LA TEMPERATURA SOBRE EL CORAZÓN
El aumento de la temperatura, como el que ocurre cuando existe fiebre, causa un gran incremento de la frecuencia cardíaca, a veces hasta el doble de la normal. La disminución de la temperatura produce grandes descensos de la frecuencia cardíaca, cayendo a tan sólo unos pocos latidos por minuto cuando la persona se encuentra cera de la muerte por hipotermia, a temperaturas de 15.5 a 21.1ºC (60 a 70 ºF).
FISIOLOGÍA DEL MÚSCULO CARDIACO
El corazón esta formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto que la duración de la contracción es mucho mayor. Por el contrario, las fibras especializadas de excitación y conducciones contraen solo débilmente porque contienen pocas fibrillas contráctiles, en cambio, presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón, formando así un sistema excitado que controla el latido rítmico cardiaco.
Músculo cardiaco como sincitio.
Las zonas oscuras que atraviesan las fibras musculares cardíacas se denominan discos discos intercalados; realmente son membranas celulares que separan las células musculares cardiacas individuales entre si. Es decir, las fibras musculares cardiacas están formadas por muchas células individuales conectadas entre si en serie y en paralelo.
En cada uno de los discos intercalados las membranas celulares se fusionan entre si de tal manera que forman uniones “comunicantes” (en hendidura) permeables que permiten una difusión de iones casi totalmente libre. Por tanto, desde un punto de vista funcional los iones se mueven con facilidad en el líquido intracelular a lo largo del eje longitudinal de las fibras musculares cardíacas, de modo que los potenciales de acción viajan fácilmente desde una célula muscular cardiaca a la siguiente, a través de los discos intercalados. Por tanto, el músculo cardiaco es un sincitio de muchas células musculares cardiacas en el que las células están tan interconectadas entre si que cuando una de ellas se excita el potencial de acción se propaga a todas, propagándose de una célula a otra a través de las interconexiones en enrejado.
El corazón realmente esta formado por dos sincitios: el sincitio auricular, que forma las paredes de las dos aurículas, y el sincitio ventricular, que forma las paredes de los dos ventrículos. Las aurículas están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV) entre las aurículas y los ventrículos. Normalmente los potenciales no se conducen desde el sincitio auricular hasta el sincitio hacia el sincitio ventricular directamente a través de este tejido fibroso. Por el contrario, sólo son conducidos por medio de un sistema de conducción especializado denominado haz AV, que es un fascículo de fibras de conducción de varios milímetros de diámetro.
Esta división del músculo del corazón en dos sincitios funcionales permite que las aurículas se contraigan un pequeño intervalo antes de la contracción ventricular, lo que es importante para la eficacia del bombeo del corazón.
PROPIEDADES FISIOLOGICAS DEL MUSCULO CARDIACO
-
Automatismo
-
Ley del todo o nada
-
Sacudida simple y tétanos
II.-Potenciales de acción en el músculo cardíaco
El potencial de acción que se registra en una fibra muscular ventricular, es en promedio de aproximadamente 105 mV. Lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente -85mV. entre los latidos hasta un valor ligeramente positivo, de aproximadamente + 20mV. Durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 segundos, mostrando una meseta seguida al final de una meseta de repolarización súbita. La presencia de esta meseta del potencial de acción hace que la contracción ventricular dure hasta 15 veces más en el músculo cardiaco que en el mundo esquelético.
¿Qué produce el potencial de acción prolongado y la meseta?
En este punto debemos preguntarnos por que el potencial de acción del músculo cardiaco es tan prolongado y por qué tiene una meseta, mientras que el del músculo esquelético no la tiene.
Al menos dos diferencias importantes entre las propiedades de la membrana del músculo cardiaco y esquelético son responsables del potencial de acción prolongado y por qué tiene una meseta. Mientras que el del músculo esquelético está producido casi por completo por la apertura súbita de grandes números de los denominados canales rápidos de sodio que permiten que grandes cantidades de iones sodio entren en la fibra muscular esquelética desde el líquido extracelular. A estos canales se les denominan canales “rápidos” porque permanecen abiertos solo algunas milésimas de segundo y después se cierran súbitamente, al final de este cierre se produce la repolarización y el potencial de acción ha terminado en otra milésima de segundo aproximadamente.
En el músculo cardiaco, el potencial de acción esta producido por la apertura de dos tipos de canales:
Los mismos canales rápidos de sodio que en el músculo esquelético
Otra población totalmente distinta de canales lentos de calcio, que también se denominan canales de calcio sólido.
Esta segunda población de canales difiere de los canales rápidos de sodio en que se abren con mayor lentitud y, lo que es incluso mas importante, permanecen abiertos durante varias décimas de segundo. Durante este tiempo fluye una gran cantidad de iones tanto calcio como sodio a través de estos canales hacia el interior de la fibra muscular cardiaca, y esto mantiene un periodo prolongado de despolarización, dando lugar a la meseta del potencial de acción. Además, los iones calcio que entran durante esta fase de meseta activan el proceso contráctil del músculo, mientras que los iones calcio que producen la contracción del músculo esquelético proceden del retículo sarcoplásmico intracelular.
La segunda diferencia funcional importante entre el músculo cardiaco y el músculo esquelético que ayuda a explicar tanto el potencial de acción prolongado como su meseta es éste: inmediatamente después del inicio del potencial de acción, la permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye aproximadamente cinco veces, un efecto que no aparece en el músculo esquelético. Esta disminución de la permeabilidad al potasio se puede deber al exceso de flujo de entrada de calcio a través de los canales de calcio que se acaba de señalar. Independientemente de la causa, la disminución de la permeabilidad al potasio reduce mucho el flujo de salida de iones potasio de carga positiva durante la meseta del potencial de acción a su nivel de reposo. Cuando los canales lentos de calcio-sodio se cierran después de 0.2 a 0.3 segundos y se interrumpe el flujo de entrada de iones calcio y sodio, también aumenta rápidamente la permeabilidad de la membrana a los iones potasio; esta rápida perdida de potasio desde la fibra inmediatamente devuelve el potencial de membrana a su nivel de reposo, finalizando de esta manera el potencial de acción.
Velocidad de la conducción de las señales en el músculo cardiaco
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de aproximadamente 0,3 a 0,5 m/s, o aproximadamente1/250 de la velocidad en la fibras nerviosas grandes y aproximadamente 1/10 de la velocidad de las fibras musculares esqueléticas. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras del Purkinje, es de hasta 4m/s en la mayoría de las partes del sistema, lo que permite una conducción razonablemente rápida de la señal excitadora hacia las diferentes partes del corazón.
Período refractario del músculo cardiaco
El músculo cardiaco, al igual que todos los tejidos excitables, es refractario a la reestipulación durante el potencial de acción. Por tanto, el periodo refractario del corazón es el intervalo de tiempo, durante en cual un impulso cardiaco normal no puede reexcitar una zona ya excitada de músculo cardiaco. El período refractario normal del ventrículo es de 0.25 a 0.30 segundos, que es aproximadamente la duración del potencial de acción en meseta prolongado. Hay un periodo refractario relativo adicional de aproximadamente 0.05 segundos, durante el cual es mas difícil de lo normal excitar el músculo pero, sin embargo, se puede excitar con una señal excitadora muy intensa, como se demuestra por la contracción “prematura” temprana.
El periodo refractario del músculo auricular es mucho más corto que el de los ventrículos (aproximadamente 0,15 segundos para las aurículas en comparación con 0,25 a 0,30 segundos para los ventrículos).
Acoplamiento excitación-contracción: función d elos iones calcio y d elos túmulos transversos
El termino “acoplamiento excitación -contracción” se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas del músculo se contraigan. Una vez más hay diferencias en este mecanismo en el músculo cardiaco que tiene efectos importantes sobre las características de su contracción.
Al igual que en el músculo esquelético, cuando un potencial de acción pasa sobre la membrana del músculo cardiaco el potencial de acción se propaga hacia el interior de la fibra muscular cardiaca a lo largo de las membranas de los túbulos transversos (T). Los potenciales de acción de los túbulos T, a su vez, actúan sobre la membrana de los túbulos sarcoplásmicos longitudinales para producir la liberación de iones calcio hacia l sarcoplasma muscular desde el retículo sarcoplásmico. En algunas milésimas de segundo más estos iones calcio difunden hacia las miofibrillas y catalizan las reacciones químicas que favorecen el deslizamiento de los filamentos de actina y de miosina entre si, lo que da lugar a la contracción muscular.
Hasta ahora este mecanismo de acoplamiento excitación-contracción es el mismo que el del músculo esquelético, aunque hay un segundo efecto que es bastante diferente. Además de los iones calcio que se liberan hacia el sarcoplasma desde los propios túbulos T en el momento del potencial de acción. De hecho, sin este calcio adicional procedente de los túbulos T la fuerza de la contracción del músculo cardiaco se reduciría de manera considerable porque el retículo sarcoplásmico del músculo cardiaco esta peor desarrollado que el del músculo esquelético y no almacena suficiente calcio para generar una contracción completa. Por el contrario, los túbulos T del músculo cardiaco tiene un diámetro cinco veces mayor que los túbulos del músculo esquelético, lo que significa un volumen 25 veces mayor. Además, en el interior de los túbulos T hay una gran cantidad de mucopolisacáridos que tienen carga negativa y que se unen a una abundante reserva de iones calcio, manteniéndolos siempre disponibles para su difusión hacia el interior de la fibra muscular cardiaca cuando aparece un potencial de acción en un túbulo T.
La fuerza de la contracción del músculo cardiaco depende en gran medida de la concentración de iones calcio en los líquidos extracelulares. La razón de esto es que las aberturas de los túbulos T atraviesas directamente la membrana de la célula muscular cardiaca hacia los espacios extracelulares que rodean las células muscular cardiaca hacia los espacios extracelulares que rodean las células, lo que permite que el mismo liquido extracelular que esta en el intersticio del músculo cardiaco se introduzca también en los túbulos T (es decir, la disponibilidad de los iones calcio para producir la contracción del músculo cardiaco) depende en gran medida de a concentración de iones calcio, en el líquido extracelular.
(A modo de contraste, la fuerza de la contracción del músculo esquelético apenas se ve afectada por cambios moderados de la concentración del calcio en el líquido extracelular porque la concentración del músculo esquelético está producida casi por completo por los iones calcio que son liberados por el retículo sarcoplásmico del interior de la propia fibra muscular esquelética.)
Al final de la meseta del potencial de acción cardiaco se interrumpe súbitamente el flujo de entrada de iones calcio hacia el interior de la fibra muscular y los iones calcio del sarcoplasma se bombean rápidamente hacia el exterior de lasa fibras musculares, hacia el retículo sarcoplásmico y hacia el espacio de los túbulos T-líquido extracelular. En consecuencia, se interrumpe la contracción hasta que llega un nuevo potencial de acción.
Duración de la contracción
El músculo cardiaco comienza a contraerse algunos milisegundos después de la llegada del potencial de acción y sigue contrayéndose hasta algunos milisegundos después de que finalice. Por tanto, la duración de la contracción del músculo cardiaco depende principalmente de la duración del potencial de acción, incluyendo la meseta, aproximadamente 0,2 segundos en el músculo auricular y 0,3 segundos en el músculo ventricular.
III.- Derivaciones
Son las diferentes posiciones de donde se observa la actividad eléctrica del corazón.
Las derivaciones se forman por la combinación entre los electrodos que se sitúan en la superficie del cuerpo.
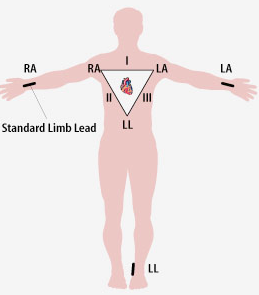
a) Derivaciones Estándar
Llamadas bipolares porque están compuestas por dos electrodos, uno negativo y otro positivo, registrándose en el trazo del electrocardiograma la diferencia de potenciales entre ambos.
D1
Se considera el brazo izquierdo como positivo y el derecho como negativo.
D2
Se considera pierna izda. positiva y brazo dcho negativo.
D3
Se considera pierna izquierda positiva y brazo izqdo negativo.
Ley de Einthoven
La Ley de Einthoven dice que el potencial de II debe ser igual a la suma de los potenciales de
I y III, en caso de no cumplirse estaríamos ante una mala colocación de los electrodos.
Se pueden desplazar los ejes de las derivaciones de los miembros al centro del triángulo que
forman, obteniendo un sistema de referencia hexaaxial, quedando separado cada eje 30º del
contiguo, permitiendo dar una orientación espacial del vector resultante de la actividad
eléctrica del corazón.
Triangulo de Einthoven
b) Derivaciones unipolares
b.1) Aumentadas
La característica general de las 3 derivaciones unipolares de miembros es su obtención a partir de un
electrodo explorador, que tiene como polo contrario (para establecer el necesario dipolo) un potencial que
no es exactamente igual a cero, pero que se aproxima mucho a esa magnitud, por lo que su fuerza es desdeñable.
Las derivaciones aumentadas se consideran unipolares porque abarcan un electrodo positivo en el brazo izquierdo, brazo derecho y pierna izquierda, que registra el potencial eléctrico en ese único punto en referencia a las otras dos derivaciones.
b.1.1) Derivación AVR
voltaje aumentado del brazo derecho. este es el electrodo positivo en referencia al brazo izquierdo y pierna izquierda.
Esta derivación mira la actividad eléctrica del corazón desde la dirección del brazo derecho.
b.1.2) Derivación AVL
voltaje aumentado del brazo izquierdo, este es el electrodo positivo en referencia al brazo derecho y pierna izquierda.
Esta derivación mira la actividad eléctrica del corazón desde la dirección del brazo izquierdo.
b.1.3) Derivación AVF
voltaje aumentado del pie izquierdo, este el electrodo positivo en referencia al brazo derecho y brazo izquierdo
Esta derivación mira la actividad eléctrica del corazón desde la dirección de la parte de abajo del corazón.
b.2) Derivaciones precordiales:
Son todas monopolares. Van de V1 a V6.
V1: 4º espacio intercostal, línea paraesternal derecha.
V2: 4º espacio intercostal, línea paraesternal izquierda.
V3: mitad de distancia entre V2 y V4
V4: 5º espacio intercostal, línea medioclavicular.
V5: 5º espacio intercostal, línea axilar anterior
V6: 5º espacio intercostal, línea axilar media.
También se pueden registrar las mismas derivaciones precordiales en el lado derecho (casos
especiales) nombrándose V3R, V4R, V5R, V6R.
III. REGISTRO
-
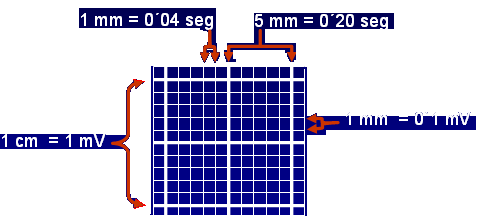
Milimetrado (Cuadriculado)
-
Cada 5 rayitas finas una
gruesa y cada 5 gruesas
una marca (1 segundo)
-
Calibrado el electrocardiógrafo para que:
-
Velocidad del papel: 25 mm/s: 1 mm de ancho = 0´04 s
-
1 cm. de altura = 1 mV 1 mm de altura = 0`1 mV
IV.-Excitación rítmica del corazón
El corazón esta dotado de un sistema especial para:
1.- generar impulsos eléctricos rítmicos para producir la contracción rítmica del músculo cardiaco
2.- conducir estos estímulos rápidamente por todo el corazón.
Este sistema también es importante porque permite que todas las porciones de los ventrículos se contraigan casi casi simultáneamente, lo que es esencial para una generaron de presión mas eficaz en las cavidades ventriculares.
SISTEMA DE EXITACION ESPECIALIZADO Y CONDUCCION DEL CORAZON
1.-El nodulo sinusal tambien denominado nodulo sinoauricular en el que se genra el impulso ritmico normal ; las vias internodales que conducen el impulso desde el nodulo sinusal hasta el nodulo auriculo ventricular en le cual el impulso originado en las auriclas se retrasa antes de penetrar en los ventrículos el haz auricloventricular que conduce el impulso desde las aurículas hacia los ventrículos y las ramas izquierda y derecha del haz de fibras de purkinge que conducen el impulso cardiaco por todo el tejido de los ventrículos.
está situado en el techo de la aurícula derecha , en el "surco terminal", ligeramente lateral a la unión de la orejuela derecha y la vena cava superior.
Las fibras de este nodulo casi no tiene filamentos musculares contráctiles sin embargo las fibras del noculo sinusal se conectan directamente con las fibras musculares de la uricula llevando asi los potenciales de accion .
2.-La Conducción a nivel Auricular( conducción internodal ) :.A nivel auricular no hay un "verdadero" tejido de conducción ( a diferencia de los ventrículos que sí cuentan con tejido de conducción : Haz de His y fibras de Purkinje ).El impulso desde el nodo sinusal al nodo AV se transmite de forma radial y sincitial (la forma más rápida posible) , de manera que se admite que hay 3 áreas de fibras musculares de conducción más rápida, que han sido llamadas : 1)."vía internodal anterior" de Bachmann 2). "vía internodal media" de Wenckebach . 3). "vía internodal posterior" de Thorel .
Esto dura 0.03s
3.-. El Nodo Aurículo-Ventricular : está localizado en la aurícula derecha , en la parte baja a la derecha del septo interauricular, en el lado derecho del anillo fibrosos central.Está localizado dentro del "triángulo de Koch", anterior al seno coronario y encima de la valva septal de la tricúspide.
El sistema de conducción auricular esta organizado de modo que el impulso cardiaco no viaja desde las auriclas hacia los ventrículos rapidamente este retraso da tiempo para que als auriculas vacien su sangre hacia los entriculos antes de que inicie la contraccio n ventricular .este retraso se debe al nodulo auriculoventricular.Esto dura 0.09s.
Se roduce un retraso final de otros 0.04 s principalmene en este haz AV penetrante que esta formad por multiples fascículos peuqeños que atraviesan el tejido fibroso que separa als auriclas d elos ventrículos .
Asid ecimos que el retraso total de la señal exitadora desde el nodo sinusal hasta el nodulo AV q hace que haya unretraso total de 0.16 contando con los 0.03 iniciales tiempo que dura las fibras internodales. Esta transmisión se dice que es lenta .
4.- El Haz de His ohaz auriculoventricular: cuando las fibras de conducción atraviesan el anillo fibroso central se designan como "haz penetrante" o "Haz de His". Cruza el trígono fibroso pasando por la parte posteroinferior del septo membranoso y se dirige en dirección anterior y medial. Tiene una longitud aproximada de 1 cm. antes de dividirse , siendo el "haz no ramificado" ( que cursa a lo largo del septo membranoso ventricular ) ( en el 75% de los casos , este "haz no ramificado" va a lo largo de la cara izquierda del septo interventricular )
5.- Ramas y Fibras de Purkinje : La porción ramificada , comienza con las fibras que forman la rama izquierda en "cascada".La rama izquierda se divide en 2 ramas principales : 1). rama antero-superior : que se dirige hacia arriba y adelante , terminando en el músculo papilar anterior. 2). rama postero-inferior: que se dirige hacia atrás y abajo, terminando en el músculo papilar posterior.
Después de dar las ramas para formar "la rama izquierda" el haz continúa como "rama derecha". Tanto la rama izquierda como derecha están "aisladas" durante su trayecto "no ramificado" , para finalmente ramificares en fibras de Purkinje que conectarán con el endocardio ventricular.
Desde el momento en que el impulso cardiaco entre las ramas de haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de purkinge el tempo total transcurrido es en promedio de solo 0.03 por tanto una vez que el impulso cardiaco ha entrado en el sistema de conducción ventricular de purkinje se propaga casi inmediatamente a toda la masa del músculo ventricular .
El músculo cardiaco envuelve el corazón en una doble espiral con tabiques fibrosos entre las capas en espiral por tanto el impulso cardiaco no viaja directamente al exterior hacia la superficie del corazón sino que se angula hacia la superficie a lo largo de las direcciones de las espirales.
V.-ELECTROCARDIOGRAMA NORMAL.
El electrocardiograma normal esta formado por una onda P un complejo QRSy una onda T. confrecuencia aunque aunque no siempre el complejo QRSesta formado por tres ondas separadas la onda, la onda R, y la onsa S.
a.-ONDAS DE DESPOLARIZACION
*. onda P .-Producida por la "despolarización" ( activación ) auricular.El inicio de la despolarización auricular es en la parte alta de la aurícula derecha y la última zona en despolarizarse es la parte distal de la aurícula izquierda ( que ocurre a los 60 miliseg (0,06 sg) del inicio de la activación auricular , por lo que la "duración" aproximada de la onda P es de 0,06 seg. )esto antes del comienzo de la contracción auricular.
*. intervalo PR : es el espacio medido entre el "inicio" de la onda P y el "inicio" del QRS . Está producido por el retraso del impulso que viene de las aurículas al pasar por el nodo A-V y en menor medida al retraso que se produce al pasar el impulso por el Haz de His.esta formado por los potenciales que se generan cuando se despolariza los ventrículos antes de su contracción es decir a medida que la onda de despolarizacion se porpaga por los ventrículos
Varía con la edad y con la frecuencia cardíaca ( puede variar desde 0,08 seg. hasta 0,18 seg.)
complejo QRS .-Está producido por la despolarización ( activación ) ventricular esta producido por los potenciales que se generan cuando los ventrículos se recuperan del estado de despolarizacion .este proceso normalmente aparece en el músculo ventricular entre 0.25 y 0.35 segundos después de la despolarizacion y la onda T .
La duración del QRS es variable , aumenta con la edad y pasa de un valor medio de 0,06 seg. en el RN a 0.08 seg. a los 12 años . Valores > de 0,10 seg. son considerados anormales. Cuando la masa ventricular aumenta , la duración del QRS aumenta.
-
Corresponde a la desporalización ventricular.
-
El voltaje del QRS es muy variable.
-
Si se produce un retraso o una interrupción de la conducción en cualquiera de las ramas del haz, el QRS se ensanchará de la manera característica del bloqueo de la rama derecha o izquierda del haz.
B.-ONDA DE REPOLARIZACION
*. onda T : está generada por la repolarización del miocardio ventricular.Es normalmente asimétrica , con un ascenso más gradual que el descenso.eneralmente tiene forma "curvada" pero puede tener una pequeña "giba"
Su amplitud generalmente no se mide ya que es muy variable , sin embargo ondas T de bajos voltajes o planas en varias derivaciones pueden indicar una anormalidad.
La repolarización es un proceso opuesto a la despolarización , pero como se realiza de epicardio a endocardio , al final el vector resultante de la onda T es paralelo al del QRS.
C.-Ondas de despolarización frente a ondas de repolarizacion
Una fibra muscular tiene cuatro fases de despolarización y repolarizacion .durante la despolarizacion el potencial negativo normal del interior de la fibra se invierte y ligeramente positivo en el interior y negativo al exterior .obsérvese también que no se registra ningún potencial en ele electrocardiograma cuando el músculo ventricular esta completamente polarizado o completamente despolarizado.Solo cuando el músculo esta parcialmente polarizado o parcialmente despolarizado hay flujo de corriente desde una parte de los ventrículos hacia la otra y por tanto la corriente también fluye hacia la superficie del cuerpo para generar electrocardiograma.
D.-Relación de la contracción auricular y vetriclar con las ondas del electrocardiograma
Antes de que pueda producir la contracción del músculo la despolarización se debe propagar por todo el músculo para iniciar los procesos químicos de la contracción ..la onda P se produce al inicio de la contracción auricular y el complejo QRS de ondas se produce al comienzo de la contracción de los ventrículos . Los ventrículos siguen contraídos hasta después de que se haya producido la repolarizacion es decir hasta después del final de la onda T .
Las aurículas se repolarizan aproximadamente 0.15 a 0.2 segundos después de la finalización de la onda P .Este momento coincide coincide aproximadamente con el momento en que se registra el complejo QRS en el electrocardiograma .Por tanto la onda de repolarizacion auricular conocida como onda T auricular habitualmente esta oscurecida por el complejo QRS que es mucho mayor .Por este motivo raras veces se observa la onda T auricular del electrocardiograma la onda de repolarizacion ventricular es la onda T del electrocardiograma normal .habitualmente el músculo ventricular comienza a repolarizarce en algunas fibras aproximadamente 0.2 segundos después del comienzo de la onda de despolarización ( el complejo QRS) pero en muchas oras fibras tarda 0.35 segundos .Así el proceso de repolarizacion ventricular se extiende a lo largo de un periodo prolongado de aproximadamente 0.15 segundos por este motivo la onda T del electrocardiograma normal es una onda prolongada aunque el voltaje de la onda T es mucho menor que voltaje del complejo QRS en parte a la duración prolongada ..
Intervalo PQ o PR entre el comienzo de la onda P y el comienzo del complejo QRS es el intervalo que hay entre el inicio de la excitación eléctrica de las aurículas y el inicio de la excitación de los ventrículos .Este periodo se llama PQ normal es de aproximadamente de 0.16 segundos.
Intervalo QT.- la contracción del ventrículo dura casi desde el comienzo de la onda Q (Rsi no hay Q) hasta el final de la onda T este intervalo se denomina intervalo Q-T y habitualmente es de aproximadamente 0.35 segundos.
A la hora de "leer" ó interpretar un electrocardiograma (ECG) debemos de tener en cuenta que hay una serie de " Medidas básicas" que siempre deberán ser obtenidas.Existen diferentes métodos o "secuencias" para obtenerlas , nosotros proponemos una secuencia de "lectura del ECG" siguiendo la cual detectaremos prácticamente cualquier alteración electrocardiográfica.
La secuencia que proponemos a la hora de la lectura de un Electrocardiograma es la siguiente
1. Frecuencia Cardiaca.
2. Ritmo
3. Eje del complejo QRS en el plano frontal.
4. Intervalos : - intervalo PR
- duración del QRS
- intervalo QTc
5. Hipertrofias : - Auriculares
- Ventriculares
6. Repolarización ( onda T ) y segmento ST
E.-MEDICION DE LA FRECUENCICA CARDIACA
L a frecencia cardiaca es el reciproco del intervalo de tiempo entre dos altidos cadiacos sucesivos el intervalo normal entre dos compleos QRS sucesivos en unapersona adulta es d aproximadamente 0.83 segundos .Esto corresponde a una frecuencia cardiaca de 60/0.83 veces por minuto o 72 latidos por minuto.
Universidad Nacional del Altiplano Facultad de Medicina
1
DEDICATORIA
A Dios, por guiar nuestro camino y estar siempre a nuestro lado. A nuestros padres por su amor, comprensión y apoyo incondicional.
AGRADECIMIENTO
A los docentes de la facultad de medicina por sus enseñanzas que ayudan a la formación de nuestra futura profesión a nuestra familia y a todos nuestros compañeros.
AGRADECIMIENTO
A los docentes de la universidad por brindarnos sus enseñanzas para una mejor formación a nuestra familia por su apoyo incondicional.
Línea axilar anterior
Línea medioclavicular
Línea axilar media
Descargar
| Enviado por: | Jessica Pilar Zapata Lipe |
| Idioma: | castellano |
| País: | Perú |
Todos los derechos reservados.