Biología
Sistemática filogenética
MÉTODOS
Y TÉCNICAS EXPERIMENTALES EN BIOLOGÍA ANIMAL
----------0----------
CURSO INTERACTIVO
DE SISTEMÁTICA FILOGENÉTICA
Indice de temas
INDICE 3
Tema 1.- Introducción a la Sistemática. La Sistemática preevolutiva. La Nueva Sistemática. El Feneticismo. 5
Problemas 11
Tema 2.- Los principios teóricos de la Sistemática Filogenética. 13
Problemas 17
Tema 3.- Los métodos de la Sistemática Filogenética. La inferencia filogenética. Caracteres apomorfos y plesiomorfos. Determinación de la polaridad de los caracteres. 19
Problemas 23
Tema 4.- Construcción de cladogramas. Ejemplos prácticos. 25
Problemas 31
Tema 5.- Cladogramas y clasificación. 33
Problemas 39
Tema 6.- Aplicaciones de la Sistemática Filogenética. Introducción a la Filogenia Molecular. 41
Problemas 45
DICCIONARIO 47
CRÉDITOS 51
Tema 1.- Introducción a la Sistemática. La Sistemática preevolutiva. La Nueva Sistemática. El Feneticismo.

La práctica de la Sistemática consiste en la nominación y ordenamiento de los seres vivos de acuerdo con unas normas aceptadas por toda la comunidad científica. En la actualidad, estas normas constituyen un sistema denominado linneano en honor de Karl Von Linneo (1707-1778). Dicho sistema se basa en dos principios:
Cada especie de ser vivo (animal, planta, protista o bacteria) recibe un nombre único, en latín, construido con dos palabras (un binomio). Dicho binomio consta de un nombre genérico, que puede ser común a varias especies próximas, y un nombre específico.Por ejemplo: Homo sapiens, Canis familiaris o Pinus pinea
La agrupación de las especies se hace en estructuras jerárquicas, de mayor o menor categoría. Esto quiere decir que los taxones (unidades de agrupación) de una categoría determinada se agrupan en taxones de superior categoría y comprenden taxones de categoría inferior. En el caso de la especie humana, esto se reflejaría en la siguiente tabla, que recoge las categorías linneanas "clásicas":
| Categoría: | Taxón: | Otros taxones próximos con la misma categoría |
| Reino | Animales (Metazoa) | Plantas, Protistas... |
| Phylum | Cordados | Equinodermos, Hemicordados... |
| Clase | Mamíferos | Arcosaurios, Anfibios... |
| Orden | Primates | Quirópteros, Dermápteros... |
| Familia | Homínidos | Póngidos, Cercopitécidos... |
| Género | Homo | Australopithecus... |
| Especie | Homo sapiens | Homo erectus |
El esquema formal de Linneo permitió poner fin a una situación bastante caótica en la organización de la información sistemática. Desde los tiempos de Aristóteles sólo se distinguía entre animales con sangre (vertebrados) y sin sangre (invertebrados), y dentro de cada grupo los animales se clasificaban en función de su modo de vida (natatilia: nadadores, volatilia: voladores, gressibilia: .... vermes:gusanos). Dentro de cada uno de estos grupos la ordenación era alfabética. Esto llevaba a situaciones tan pintorescas como la de colocar uno junto a otro, en los libros de animales del Renacimiento, al caballito de mar (Hippocampus en latín) con el hipopótamo, ya que ambos viven en el agua y sus nombres empiezan por "hip".
Linneo, por tanto, proporciona un sistema formal de agrupación con una gran capacidad para almacenar información. Pero es preciso preguntarse ahora: ¿Cuál es el criterio del naturalista para agrupar determinados taxones (p.e. especies o familias) dentro de un taxón de categoría superior (p.e. géneros u órdenes)? Cuando a finales del XIX se impuso la idea de evolución,se pudo utilizar con este fin el parentesco filogenético, como luego veremos. Pero, ¿cómo se hacían las agrupaciones antes de este acontecimiento?
El criterio fundamental también lo proporciona Aristóteles, y se basa en el esencialismo. El naturalista trata de distinguir, en las características de los organismos, las que son esenciales (las que definen su "esencia", su "causa formal") de las que no lo son (caracteres accidentales, no esenciales). De esta forma una especie determinada, un género o una familia concreta revelan caracteres "diagnósticos". Estos caracteres diagnósticos, básicamente morfológicos, permiten agrupar a los individuos en especies, o a los taxones de inferior categoría en taxones de categoría superior. La tarea del sistemático sería, por tanto, la de identificar los caracteres esenciales y realizar agrupaciones (y clasificaciones) en función de ellos.
Así, decimos de este organismo que es un tiburón (técnicamente: pertenece a los elasmobranquios galeomorfos) porque reúne las características esenciales de los elasmobranquios (endoesqueleto cartilaginoso, dientes de sustitución, escamas placoideas) y las de los galeomorfos (cinco hendiduras branquiales y aleta anal). Por supuesto, cualquier otro ser vivo que reúna los cinco caracteres esenciales antes citados debe ser considerado tiburón (elasmobranquio galeomorfo). Por otro lado, este individuo tiene una serie de caracteres, como la aleta caudal semilunar o el color gris, que no influyen en la consideración de que se trata de un tiburón, aunque pueden ser caracteres esenciales a la hora de determinar el género o la especie.
En este contexto esencialista, de raíz aristotélica, es donde podemos situar el concepto tipológico de especie. En efecto, si decimos que un animal concreto, un individuo, pertenece a la especie Canis familiaris (el perro) es porque reúne una serie de características esenciales para ser considerado como tal (junto con otras no esenciales que hacen de dicho individuo un caniche, un pequinés o un chihuahua).
Este criterio, claro y sencillo, se complicó cuando progresivamente, a lo largo del XIX, se fue imponiendo la idea de evolución. En efecto, si todos los seres vivos son el resultado de un proceso evolutivo, contamos con una nueva variable a la hora de realizar agrupaciones: el parentesco filogenético. En el próximo tema desarrollaremos esta noción. Bástenos ahora señalar que, en efecto, hoy en día existen dos criterios para realizar agrupaciones de taxones:
La similitud morfológica, es decir, la compartición de características definitorias del taxón. Agrupamos a las aves en el taxón Aves porque tienen plumas, alas, pico córneo, etc..., es decir porque reúnen los caracteres "esenciales" de las aves.
El parentesco filogenético, es decir, la proximidad de los ancestrales comunes. Agruparemos a las aves en un mismo taxón porque todas comparten un ancestral común relativamente reciente y, desde luego, más reciente que el ancestral que cualquier ave comparte con cualquier otro vertebrado.
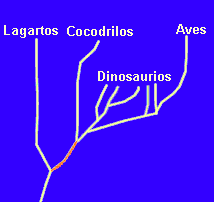
En el caso anterior no hay conflicto. Las aves estarán agrupadas en el taxón Aves independientemente del criterio elegido. Pero en realidad el conflicto entre ambos criterios puede existir. Un ejemplo muy típico es el de la agrupación de lagartos, cocodrilos y aves. Si atendemos a criterios morfológicos, a primera vista lagartos y cocodrilos se parecen más entre sí, lo que ha justificado su agrupación durante siglos en el taxón "Reptiles". En cambio, la filogenia del grupo (representada en la figura) señala que aves y cocodrilos son ramas derivadas de un linaje común (señalado en naranja) y por tanto, como veremos en el próximo tema, comparten un mayor grado de parentesco. Dicho parentesco favorece una agrupación Cocodrilos+Aves en un taxón que se ha denominado "Arcosaurios". Luego veremos las consecuencias de este tipo de conflictos
Recuerda, podemos agrupar a los organismos en función de sus semejanzas morfológicas o en función de su parentesco filogenético.
La Nueva Sistemática o Sistemática Evolutiva, una corriente desarrollada a partir de los años 40 por Ernst Mayr y Georges Simpson, constató que desde la aceptación de la idea de la evolución no se habia producido ningún cambio en la práctica de la Sistemática. Es decir, se seguía trabajando como en los tiempos de Linneo. La Sistemática Evolutiva propuso una serie de mejoras metodológicas, tales como:
| | |||
|
| La utilización del concepto biológico de especie, basado en propiedades biológicas (la comunidad reproductiva) y no el concepto tipológico, basado en en el esencialismo, como vimos antes. | ||
|
| La utilización de caracteres morfológicos y no morfológicos, y la necesidad de valorar adecuadamente (con métodos estadísticos si es necesario) la variabilidad. | ||
|
| La necesidad de ponderar similitud morfológica y parentesco filogenético en caso de conflicto. | ||
Estas propuestas fueron bastante criticadas a lo largo de los años 60. Por ejemplo, se señaló que el concepto biológico de especie no era operativo. En la práctica no se comprueba, en la mayoría de los casos, el criterio de interfecundidad. Lo que es peor, especies claramente diferentes, y que no se cruzan en la naturaleza, hibridan sin problemas en condiciones de laboratorio. En realidad, ante las dificultades de la aplicación del concepto biológico de especie, los taxónomos siguen usando criterios tipológicos a la hora de definirlas.
Por otro lado, no quedaba claro cómo había que proceder en caso de conflicto entre similitud y parentesco. También se señaló que los criterios de agrupación en categorías supraespecíficas eran subjetivos. Que una familia determinada comprendiera dos, tres o cuatro géneros no podía justificarse de forma objetiva, y la aceptación de una u otra posibilidad parecía depender de la autoridad del que realizaba la agrupación. De esta forma la práctica sistemática parecía quedar fuera del dominio científico.
Antes esas críticas surgieron dos alternativas que pretendían un método sistemático científico, es decir, objetivo y replicable. La primera de estas alternativas se denominó Feneticismo o Taxonomía Numérica. La segunda, que será la que expondremos a lo largo de este curso, es la Sistemática Filogenética.
Los principios de la Taxonomía Numérica fueron desarrollados por Sokal y Sneath en dos libros publicados en 1963 y 1973. La idea básica era la de resolver el dilema similitud/parentesco en beneficio de la primera. Utilizar sólo la similitud morfológica y cuantificarla en forma de Indices de Similitud o de Distancia. Dichos índices deberían reflejar el parecido global entre los taxones.
No es objetivo de este curso desarrollar los procedimientos feneticistas. Comentaremos que estos procedimientos se basan en el uso de matrices en las que se incluyen valores numéricos de determinados atributos de las unidades a agrupar, unidades llamadas OTUs (de operative taxonomical unity) que pueden ser individuos, muestras de poblaciones o especies, etc. Los atributos pueden ser medidas, proporciones, recuentos o caracteres semicuantitativos (p.e. 0=ausencia, 1=presencia). La idea es que cuanto más similares sean dos OTUs, menor será la diferencia global entre los valores de sus atributos. De esta forma se pueden definir índices de distancia fenética, como por ejemplo la media cuadrática de las diferencias entre cada par de atributos. En el caso de identidad total entre dos OTUs dicho índice valdría 0, y será mayor cuanto más disimilares sean los OTUs.
Ejemplo de matriz de datos:
| OTU1 | OTU2 | OTU3 | |
| Car. 1 | 24 | 26 | 29 |
| Car. 2 | 12,2 | 14 | 15 |
| Car. 3 | 2 | 3 | 3 |
Ejemplo de Indices de distancia:
| OTU1 | OTU2 | |
| OTU2 | 1,89 | |
| OTU3 | 3,34 | 1,49 |
La representación gráfica de los índices de distancia lleva a diagramas denominados fenogramas, que representan los grupos de OTUs en función de su similitud global. Existen varios tipos de algoritmos de representación. El ejemplo anterior se muestra en esta figura:
Los OTUs 2 y 3, con mayor parecido global, se sitúan a una distancia de 1,49. La distancia del OTU 1 al grupo (OTU2+OTU3) es igual a la media aritmética de las distancias a cada uno de sus componentes, es decir: (1,89+3,34)/2 = 2,61.
La situación en la que se enmarca el nacimiento de la Sistemática Filogenética es, por tanto, una sistemática "evolutiva" que a pesar de su nombre seguía utilizando procedimientos básicamente esencialistas o preevolutivos. El concepto biológico de especie resultaba difícil de aplicar en la práctica, por lo que se aplicaba el concepto tipológico clásico. Se planteaban conflictos irresolubles entre similitud y filogenia. Y en última instancia la clasificación contaba con un componente fuertemente subjetivo. La alternativa feneticista, por otra parte, abandonaba no ya el concepto de evolución, sino cualquier sentido biológico en la clasificación. La similitud se conviertía en la única propiedad relevante, con lo que trabajar con seres vivos no resultaba diferente a trabajar con cualquier tipo de objetos. Veremos en los próximos temas como la Sistemática Filogenética intenta resolver estos problemas.
Ejercicios del tema 1
1.- Cada especie biológica recibe un nombre formado por:
2.- Cada unidad de agrupación (y de clasificación) recibe el nombre de:
3.- Cada nivel de agrupación (y de clasificación) recibe el nombre de:
4.- En el caso del taxón aves, ¿cuáles de estos caracteres considerarías esenciales para la definición del taxón?
patas, plumas, pico, uñas, pulmones, ojos.
5.- Los criterios que pueden utilizarse para realizar agrupaciones son:
6.- El único criterio que utiliza la taxonomía numérica es el de:
Tema 2.- Los principios teóricos de la Sistemática Filogenética.
Como hemos dicho en el tema anterior, la agrupación de taxones de una categoría inferior (p.e. especies o géneros) en taxones de categoría superior (géneros o familias, respectivamente), puede hacerse siguiendo criterios de similitud y/o de parentesco filogenético. Esto, como también hemos visto, puede dar lugar a conflictos. La solución que propone la Sistemática Filogenética es utilizar exclusivamente, el parentesco filogenético como criterio de agrupación.
Vamos a desarrollar la noción de parentesco filogenético, que es crucial para comprender lo que sigue. Se puede establecer una analogía con el parentesco familiar. ¿Qué es lo que nos hace estar emparentados con nuestros hermanos, primos o tíos? El hecho de compartir ancestrales comunes. De hecho, existe un grado de parentesco que está relacionado con la "distancia" temporal a nuestros ancestrales.
Observemos la figura. Decimos que estamos más emparentados con nuestros hermanos que con nuestros tíos o primos porque con nuestros hermanos compartimos ancestrales más cercanos (nuestros padres). En cambio, los ancestrales más recientes que compartimos con nuestros primos son nuestros abuelos. Con familiares más lejanos (primos segundos o terceros, tíos abuelos, etc.) vamos compartiendo ancestrales comunes más remotos en el tiempo (bisabuelos, tatarabuelos...).
Si seguimos ascendiendo en nuestro árbol genealógico comprenderemos que estamos emparentados, en mayor o menor grado, con todos los humanos, una consecuencia directa de una propiedad biológica, la reproducción.
Del mismo modo que todos los humanos (y todos los seres vivos) están emparentados, también existe un grado de parentesco filogenético que dependerá de la distancia a los ancestrales comunes más recientes. En este sentido decimos que los cocodrilos están más emparentados con las aves que con los lagartos porque comparten con ellas un ancestral común más reciente que el que comparten con los lagartos (puedes comprobarlo en la figura del tema 1).
Esta propiedad descansa sobre dos asunciones: que el origen de la vida ha sido único y que la diversidad de los seres vivos es el producto de la evolución. Al mismo tiempo esta propiedad nos está proporcionando un criterio objetivo de agrupación. En Sistemática Filogenética agruparemos los taxones en función de su grado de parentesco filogenético, es decir, en función del orden relativo de sus ancestrales comunes.
Recuerda: El grado de parentesco filogenético es objetivo independiente del investigador. Valorar el grado de similitud, en cambio, implica un componente subjetivo inevitable.
La práctica de la Sistemática Filogenética consistirá, por tanto en dilucidar cuál es el grado de parentesco entre los taxones y realizar agrupaciones que se basen en dicho grado de parentesco. Esto nos lleva a una primera consecuencia muy importante. Dado que los taxones están compuestos por organismos emparentados (descendientes por tanto de un ancestral común a todos ellos) todos los taxones, todas las unidades de clasificación, deberán ser estrictamente monofiléticos (ver figura). Es decir, una agrupación no monofilética, no formada por todos y cada uno de los descendientes de un ancestral común, no puede ser constituida como taxón.
| Concepto de monofilia estricta. |
Esto cambia de una manera radical la Sistemática clásica, basada en relaciones de similitud. Por ejemplo, Equinodermos, Artrópodos, Aves o Mamíferos pueden ser considerados como taxones, en tanto que asumimos que son monofiléticos. Sin embargo, los peces óseos (Osteictios), los Reptiles y los Anfibios no forman grupos monofiléticos y no pueden constituirse en taxones.
Por tanto, el sistemático filogenético debe:
Definir los grupos monofiléticos con los que va a trabajar.
Inferir las relaciones filogenéticas entre ellos.
Trasladar dicho esquema de relaciones al sistema formal linneano.
Todo esto se va a hacer en un marco fuertemente hipotético. La práctica de la Sistemática Filogenética consiste básicamente en elaborar hipótesis sobre el grado de parentesco que reflejen lo que realmente sucedió en la historia evolutiva de los seres vivos. Las hipótesis se elaboran, como ya veremos, de acuerdo con ciertas normas rigurosas. Dichas hipótesis se verán progresivamente reforzadas por nuevos descubrimientos o bien serán refutadas y abandonadas por hipótesis alternativas, más sólidas. Por tanto, el avance se producirá por ensayo y error. Esto es lo que se ha llamado el método hipotético-deductivo por filósofos de la ciencia como Karl Popper. La Sistemática Filogenética, por tanto, sitúa la práctica sistemática dentro de los dominios científicos.
Ejercicios del tema 2
1.- En Sistemática Filogenética, taxones de categoría A serán agrupados en un mismo taxón de categoría superior B siempre que...
Tengan algún ancestral común.
Exista un nivel suficiente de similitud morfológica.
Estén filogenéticamente más emparentados entre sí que
con miembros de otros taxones de igual o superior
categoría.
Su ancestral común pertenezca al taxón de categoría B
2.- Diremos que A y B están más emparentados entre sí que con un tercer taxón C si:
- A y B comparten un ancestral común más reciente que el
ancestral común que ambos comparten con C.
- A y B comparten un número mayor de caracteres
morfológicos.
- A y B no comparten ancestrales comunes con C.
3.- ¿Cuántos grupos monofiléticos puedes contar en este árbol filogenético?

4.- A partir de la figura del tema 1, que reproducimos aquí, ¿podemos afirmar que el grupo formado por lagartos + cocodrilos + dinosaurios + aves es monofilético? ¿Y el grupo de los dinosaurios es monofilético? ¿Por qué?
5.- Decimos que la Sistemática Filogenética es "científica" porque:
Maneja "hipótesis de trabajo" acerca del grado de parentesco entre los organismo.
Los que la practican son científico.
Las relaciones con las que trabaja, las relaciones de parentesco, son objetivas, es decir, realmente existentes en la Naturaleza.s. s.
Tema 3.- Los métodos de la Sistemática Filogenética. La inferencia filogenética. Caracteres apomorfos y plesiomorfos. Determinación de la polaridad de los caracteres.
En los temas anteriores hemos dejado claro que la Sistemática Filogenética utiliza las relaciones de parentesco filogenético como criterio a la hora de definir los taxones y de agruparlos en taxones de categoría superior. Esto quiere decir que el proceso fundamental consiste en hacer una inferencia filogenética, es decir, elaborar una hipótesis acerca del orden de aparición de los ancestrales comunes más recientes. Veamos un ejemplo de esto.
Supongamos tres taxones cualesquiera A, B y C. Para poder trabajar, debemos suponer de entrada que son estrictamente monofiléticos, es decir, que todos y cada uno de sus componentes derivan de un ancestral común no compartido con otros taxones. Estos taxones son resultado de una historia evolutiva que no podemos conocer con detalle. Pero supongamos por un momento que sí podemos conocer cuál ha sido la historia de los linajes de A, B, y C, y que dicha historia se representa en la figura adyacente.
Volvamos a nuestro problema. Para tres taxones cualesquiera (A, B y C) pueden formularse tres y sólo hipótesis de parentesco que podemos representar de la siguiente forma (estos diagramas se denominan cladogramas, veremos luego qué son y cómo se construyen):
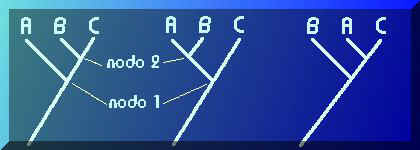
En el primer caso se propone que B y C están más emparentados porque su ancestral común (representado por el nodo 2 o unión de las dos ramas que llevan a B y C) es más reciente que el ancestral que ambos comparten con A (representado por el nodo 1). En el segundo caso los más emparentados son A y B, y en el tercer caso A y C. No hay más posibilidades.Observa que el orden de los nodos (que representan ancestrales comunes más recientes) nos indica el grado relativo de parentesco.
Si volvemos a la primera figura de este tema, la que representa lo que realmente ocurrió, nos decantaremos por la primera de las tres hipótesis. En efecto, B y C tienen toda una serie de ancestrales comunes que son más recientes que los ancestrales comunes que comparten A y B, por un lado, y A y C por el otro. Pero como no es posible conocer de forma cierta la filogenia (¡a menos que tengamos el don de la clarividencia!), debemos buscar criterios objetivos que nos permitan elegir una de las tres hipótesis.
La clave nos la va a proporcionar el análisis de los caracteres de los organismos. Consideremos un organismo cualquiera, por ejemplo, un ratón. Podemos reconocer en el ratón caracteres que han tenido un origen más o menos remoto en el tiempo. Por ejemplo, sus incisivos de crecimiento continuo, típicos de los roedores, constituyen un carácter relativamente reciente. La presencia de pelos o glándulas mamarias, características de mamíferos, deben de tener un origen mucho más antiguo que el desarrollo de dichos incisivos especiales. Aún más antigua es la presencia de cráneo o vértebras, características de los vertebrados. Y todavía más remoto en el tiempo es la presencia de uniones intercelulares especializadas o colágeno (caracteres de metazoos) o núcleo celular (eucariotas). En Sistemática Filogenética se habla de caracteres apomorfos (recientes) o plesiomorfos (antiguos)
| Caracteres son los atributos homólogos que utilizamos para establecer relaciones de parentesco (por ejemplo, número de dedos). Estados de carácter son las distintas variantes que pueden adoptar los caracteres (por ejemplo 1,2 ó 5 dedos). En la práctica, para facilitar las cosas, no haremos esta distinción en general y hablaremos de caracteres salvo que se indique lo contrario. |
De lo antes dicho se desprende que existe un orden temporal en la aparición de los caracteres o, utilizando una terminología más rigurosa, de los estados de los caracteres (ver recuadro). Como veremos a continuación, si podemos establecer un orden de aparición de determinados caracteres, si podemos distinguir entre estados ancestrales o plesiomorfos y estados avanzados o apomorfos, tendremos una herramienta para realizar inferencias filogenéticas.
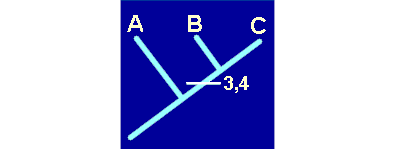
Volvamos a nuestro problema con los taxones A, B y C. En esta figura, la que representa lo que realmente sucedió, hemos expuesto la historia de algunos caracteres de los taxones. Podemos comprobar cómo los caracteres 1 y 2 se originaron en una etapa muy antigua, muy anterior a la diversificación de los linajes de A, B y C. Los caracteres 3 y 4, en cambio, aparecieron en el linaje del ancestral común de B y C. Por último los caracteres 5, 6 y 7 han aparecido en los linajes de los taxones A, B y C, respectivamente.
Si hemos entendido esto comprenderemos que el taxón A presente los caracteres 1,2 y 5. El taxón B posee los caracteres 1,2,3,4 y 6. Por último, los caracteres 1,2,3,4 y 7 estarán presentes en el taxón C.
De acuerdo con esto, el hecho de que B y C compartan dos caracteres recientes, derivados o apomorfos (3 y 4), es una consecuencia directa de su parentesco, dado que recibieron dichos caracteres de un ancestral común, no compartido con A. En este caso hablaremos de una sinapomorfía. En cambio, que A, B y C compartan caracteres primitivos o plesiomorfos como el 1 ó el 2, heredados de un lejano ancestral común de todo el grupo, no añade ninguna información. Esta compartición se denomina simplesiomorfía. Por último, tampoco nos informa de nada la posesión de caracteres derivados y no compartidos, como los 5, 6 y 7. A estos caracteres se les denomina autopomorfías.
Debe quedar claro, por tanto, que las sinapomorfías, los caracteres compartidos por dos o más taxones y derivados (aparecidos) dentro del grupo que estamos estudiando, son las que revelan relaciones significativas de parentesco.
Recuerda: Sólo los caracteres derivados y compartidos por dos o más taxones, sirven para establecer hipótesis de parentesco y se denominan SINAPOMORFIAS.
Si hemos entendido esto, el problema se va a reducir, en la práctica a:
Determinar las sinapomorfías, qué caracteres son derivados dentro del grupo y compartidos por dos o más taxones.
Construir un diagrama que refleje la hipótesis de relaciones de parentesco a partir de dichas sinapomorfías.
Estos diagramas de relaciones de parentesco, que ya hemos presentado arriba, se llaman cladogramas. Cada uno de los nodos de un cladograma representa un hipotético ancestral común para los dos taxones que parten de dicho nodo (dichos taxones que nacen de un mismo nodo son grupos-hermanos. En los cladogramas deben siempre especificarse las sinapomorfías que justifican todos y cada uno de los nodos.
Bien, pues sólo nos queda, antes de pasar a la práctica, saber qué criterios podemos utilizar para determinar la polaridad de los caracteres, es decir, el sentido del cambio, cuál es el estado ancestral y cuál el derivado.
El criterio más utilizado es el de la comparación con grupos externos (out-groups). Volvamos a la figura del principio, ampliada para incluir otro linaje próximo. Podemos ver cómo los caracteres plesiomorfos, más antiguos (1,2) aparecen en grupos relacionados con el nuestro, pero cercanos a él (G.E. en la figura). Es evidente que los estados de carácter aparecidos dentro de nuestro grupo (3,4,5,6,7) no van a ser encontrados fuera de él, en un grupo externo. ¡El carácter se "desplaza" hacia delante en el tiempo, no hacia atrás! Por tanto, el criterio nos dice que los estados plesiomorfos de carácter van a ser frecuentemente encontrados en grupos externos, mientras que los apomorfos no lo serán.
Un segundo criterio nos lo proporciona el desarrollo embrionario. En efecto, como notó el embriólogo K.E. Von Baer, los caracteres generales (plesiomorfos) aparecen siempre antes que los especiales (apomorfos). Por tanto la ontogenia de los caracteres refleja la polaridad del cambio.
Ya podemos resolver nuestro problema. Si examinamos la distribución de caracteres en los tres taxones A, B y C resolvemos:
Descartar los caracteres 1 y 2 por ser simplesiomorfías.
Descartar los caracteres 5, 6 y 7 por ser autopomorfías (derivados, pero no compartidos)
Aceptar como sinapomorfías los caracteres 3 y 4, que asumimos como derivados dentro de nuestro grupo y compartidos por los taxones B y C.
Por consiguiente, elegimos el primero de los tres cladogramas, ya que las sinapomorfías 3 y 4 revelan una posible relación de parentesco, un ancestral común a través del cual los taxones B y C recibieron los caracteres 3 y 4.
Ejercicios del tema 3
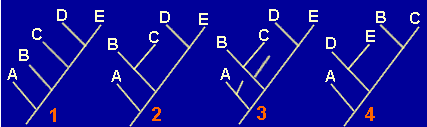
Dado el siguiente árbol filogenético:
Los números representan la aparición de los caracteres. Todos aparecen dentro del grupo excepto el carácter 1, que procede de un ancestral remoto.
1.- ¿Que caracteres encontraremos en cada uno de los taxones?
Taxón A:
Taxón B:
Taxón C:
Taxón D:
Taxón E:
2.- ¿Cuáles de los caracteres serán considerados como...
Sinapomorfías.
Simplesiomorfías.
Autopomorfías
3.- ¿Cuál de los siguientes cladogramas representa mejor la filogenia del grupo?
Tema 4.- La construcción de cladogramas. Ejemplos prácticos.
Vamos a abordar ahora casos prácticos de construcción de cladogramas, utilizando ejemplos reales. Comenzaremos por el ejemplo que utilizamos en el primer tema y que se ha convertido en un clásico a la hora de comparar las clasificaciones evolutivas y filogenéticas: las relaciones de reptiles escamosos (lagartos y serpientes), cocodrilos y aves.
Algunas de las semejanzas entre cocodrilos y escamosos (y diferencias con las aves) se recogen en esta tabla:
| Reptiles escamosos | Cocodrilos | Aves |
| Escamas córneas | Escamas córneas | Plumas |
| Patas anteriores marchadoras | Patas anteriores marchadoras | Alas |
| Mandíbulas normales | Mandíbulas normales | Mandíbulas formando un pico |
| Ectotermos | Ectotermos | Endotermos ("sangre caliente") |
Se podrían citar otras semejanzas, pero basta con estas. Un primer análisis mostrará que todas ellas, sin excepción, son simplesiomorfías (recordemos, caracteres derivados de un ancestral lejano y común a todo el grupo). El análisis de grupos externos (anfibios, tortugas...) permite encontrar sin dificultad las escamas córneas, las patas anteriores marchadoras, dientes y ectotermia. Luego, de acuerdo con lo que hemos visto acerca de las simplesiomorfías, todas estas semejanzas heredadas de ancestrales remotos no indican relación de parentesco.
En cambio sí que es posible encontrar caracteres derivados (apomorfos) y compartidos por cocodrilos y aves, es decir, sinapomorfías. En concreto:
| Dientes tecodontos (en alvéolos)* |
| Ventana preorbitaria** |
| Pelvis modificada*** |
| Tabicación completa del ventrículo cardiaco y reducción o desaparición del arco aórtico izquierdo**** |
*Presentes en las aves primitivas (Archaeopteryx). Desaparecen en las aves modernas.
**Presente en los antepasados de los cocodrilos. Desaparece en cocodrilos modernos.
***Consiste en la curvatura hacia abajo de pubis e isquion. A veces el pubis se dirige hacia atrás o desaparece.
****Existe un pequeño orificio (foramen de Panizza) en cocodrilos.
No existe ningún carácter derivado y compartido por aves y reptiles escamosos. Por ello, el cladograma que mejor refleja este análisis de caracteres es el siguiente:
En realidad sabemos que este cladograma, que hemos obtenido por medio del análisis de caracteres, se ajusta perfectamente a lo que se conoce acerca de la filogenia de los reptiles y del origen de las aves. En efecto, tanto las aves como los cocodrilos son los únicos supervivientes de determinados grupos de dinosaurios, mientras que los reptiles escamosos, aunque emparentados con dinosaurios, cocodrilos y aves, comparten ancestrales comunes más alejados en el tiempo.
¿Que consencuencias tiene para la clasificación de los vertebrados esta forma de realizar el análisis filogenético? La más evidente es que, dado que sólo podemos establecer taxones monofiléticos, no resulta aceptable un taxón Reptiles. Ha quedado claro que los reptiles no constituyen un grupo monofilético, definido por sinapomorfías. En efecto, no hay sinapomorfías que unan a cocodrilos y lagartos, como antes hemos expuesto, pero tampoco las hay entre tortugas y lagartos, por ejemplo. Sin embargo, cocodrilos, dinosaurios y aves sí pueden ser reunidos en un taxón definido por las sinapomorfías antes citadas y por otras. Dicho taxón se denomina Arcosaurios.
Recuerda: En Sistemática Filogenética sólo los grupos estrictamente monofiléticos pueden ser considerados como taxones.
Vamos a complicar un poco la cuestión, intentando insertar, en el cladograma anterior, a quelonios y mamíferos, para tener de esta forma representantes de los principales grupos actuales de amniotas. Para ello vamos a manejar la siguiente tabla de caracteres, utilizando a los anfibios como grupo externo:
| Quelonios | Mamíferos | Escamosos | Cocodrilos | Aves | Anfibios (grupo externo) | |
| Aberturas temporales en el cráneo | No tienen | Un par | Dos pares | Dos pares | Dos pares | No tienen |
| Ventana preorbitaria | Ausente | Ausente | Ausente | Presente* | Presente | Ausente |
| Regulación de la temperatura | Ectotermos | Endotermos | Ectotermos | Ectotermos | Endotermos | Ectotermos |
| Pelvis | Normal | Normal | Normal | Modificada | Modificada | Normal |
| Arco aórtico principal | Izq. y Derecho | Izquierdo | Izq. y Derecho | Derecho | Derecho | Izq. y Derecho |
*En los antepasados de los cocodrilos, como ya hemos dicho antes.
De acuerdo con el criterio del grupo externo, podemos considerar como derivados los caracteres siguientes:
Un par de aberturas temporales en el cráneo*.
Dos pares de aberturas temporales en el cráneo*.
Presencia de ventana preorbitaria
Endotermia.
Pelvis modificada.
Sólo arco aórtico izquierdo*.
Sólo arco aórtico derecho*.
*A partir del criterio del grupo externo, no podemos decidir si uno de estos caracteres es derivado respecto del otro, o si ambos son independientes. Lo que está claro es que ambos son derivados respecto de la ausencia de ventanas o del doble arco aórtico.
Son derivados y no compartidos (autopomorfías) la presencia de un par de aberturas temporales y la posesión de arco aórtico izquierdo (condiciones ambas de mamíferos). Por tanto no nos servirán para establecer relaciones de parentesco. Si utilizamos como sinapomorfías los restantes, podremos generar no uno, sino muchos cladogramas, de los que trabajaremos con estos tres:
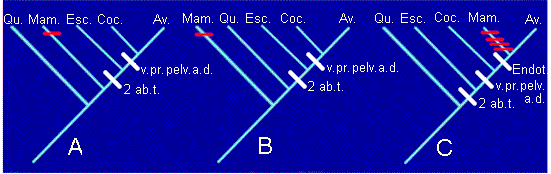
2 ab.t.= 2 aberturas temporales. v.pr.=ventana preorbitaria. pelv.=pelvis modificada. a.d.=arco aórtico derecho. Endot=endotermia.
Podemos comprobar como en los cladogramas A y B asumimos que la endotermia apareció de forma independiente en Mamíferos y Aves (convergencia). En cambio, el cladograma C asume que la endotermia estaba presente en el ancestral común de Mamíferos y Aves, pero en este caso hay que aceptar que en el linaje de los mamíferos el número de aberturas temporales pasa de dos a una, se pierde la pelvis modificada, la ventana preorbitaria y se sustituye el arco aórtico derecho por el izquierdo (reversiones). Cada uno de estos cambios (convergencias o reversiones) se representa por una raya roja en el cladograma.
¿Por cuál optaremos? En los casos en los que la distribución de caracteres en los taxones pueda explicarse por más de un cladograma (en realidad esto incluye a todos los casos posibles), aplicaremos el llamado criterio de máxima parsimonia. Este criterio aconseja a elegir aquel cladograma más sencillo, más simple, es decir, el que implique el menor grado de homoplasia. La homoplasia se puede definir como la existencia de similitud no debida a parentesco. El grado de homoplasia es el número de convergencias o reversiones que es preciso asumir a la hora de construir el cladograma. En el caso anterior, los cladogramas A y B tienen una convergencia (no dos, ¡ojo!, la convergencia se produce cuando la endotermia aparece por segunda vez). El cladograma C tiene cuatro reversiones (número de aberturas temporales, pelvis, ventana preorbitaria y arco aórtico derecho).
En conclusión, el criterio de máxima parsimonia nos haría descartar el cladograma C. Sin embargo, no podemos decidir entre el A y el B, puesto que ambos están igualmente justificado por sinapomorfías y por grado de homoplasia. Esta situación es frecuente en Sistemática Filogenética. Es necesario buscar más sinapomorfías que nos permitan decidir entre las dos opciones A y B. De hecho, si hubiéramos utilizado más caracteres hubiéramos terminado decantándonos por el cladograma B.
Vamos a utilizar el ejemplo anterior para ilustrar algunas cuestiones importantes de la Sistemática Filogenética. Por ejemplo la equivalencia potencial de convergencias y reversiones que justifica el término común de homoplasia. Observemos el siguiente cladograma:
A y B comparten un carácter apomorfo cualquiera simbolizado por el punto rojo. Sin embargo A y B no están más emparentados que B y C, que son grupos hermanos por compartir más caracteres derivados (no representados en la figura). El carácter "punto rojo" es, por tanto, un caso de homoplasia. Sin embargo, no es posible decidir entre los dos escenarios mostrados en la figura, una reversión (a la izquierda) o una convergencia (a la derecha). Cualquiera de las dos posibilidades explica la homoplasia de forma igualmente parsimoniosa.
Si hemos entendido el concepto de homoplasia, entenderemos también el concepto de "longitud del árbol" (treelength en inglés). Imaginemos cada uno de los cambios como un "paso" que añade una unidad de longitud al árbol. En el caso de nuestro cladograma anterior (elimino a los quelonios para simplificar) tenemos:
El equivalente al árbol A requiere un total de 8 cambios para explicar la distribución de los caracteres. Por tanto, la longitud del árbol será de 8 pasos. Comprueba que la hipótesis B implica un árbol de la misma longitud.
(observa que la endotermia aparece dos veces)
En cambio, el equivalente del árbol C requiere un mínimo de 10 cambios para explicar la distribución de caracteres en los taxones. Su longitud será de 10 pasos.
(la posición de los mamíferos explica la presencia de endotermia, pero a cambio es preciso asumir la pérdida de la ventana preorbitaria, la pelvis modificada, el arco aórtico derecho y un par de aberturas temporales).
La longitud del árbol nos proporciona otra forma de enunciar el criterio de parsimonia. Elegiremos aquel cladograma que implique una menor longitud, es decir, que implique el menor número de cambios. Por otro lado podemos expresar esto de forma cuantitativa. En efecto, podemos definir un índice que relacione el número mínimo de cambios posible con el número real de cambios. Esto es lo que se denomina el Indice de consistencia. En el ejemplo anterior, el número mínimo de cambios en los caracteres es de siete, los mismos que se detallan en la tabla de arriba (si no los recuerdas pulsa aquí).
Así, los árboles correspondientes a las hipótesis A y B tiene una longitud de 8 pasos sobre un mínimo de 7. El Indice de consistencia será 7/8=0,875.
El árbol correspondiente a la hipótesis C tiene una longitud de 10 pasos sobre el mínimo de 7. Su Indice de consistencia será 7/10=0,7.
Aquí tenemos otra formulación del criterio de parsimonia: Entre dos cladogramas posibles elegiremos el que presente un mayor Indice de consistencia. Podemos comprobar cómo no pudimos en su momento decidir entre las hipótesis A y B, porque ambas presentaban el mismo grado de consistencia. En cambio, la consistencia de la opción C, y de cualquier otro cladograma alternativo, siempre será inferior a 0.875.
Ejercicios del tema 4
Dada la siguiente tabla de caracteres:
| Anélidos | Artrópodos | Hemicordados | Moluscos | Cordados | Platelmintos (grupo externo) | |
| Formación de la boca | Protóstomos | Protóstomos | Deuteróstomos | Protóstomos | Deuteróstomos | Protóstomos |
| Formación del celoma | Esquizocelia | Esquizocelia | Enterocelia | Esquizocelia | Enterocelia | Sin celoma |
| Teloblastos (precursores del mesodermo) | Presentes | Presentes | Ausentes | Presentes | Ausentes | Ausentes |
| Metamería | Presente | Presente | Ausente | Ausente* | Presente** | Ausente |
| Posición del cordón nervioso | Ventral | Ventral | Dorsal | Ventral | Dorsal | Ventral |
| Hendiduras faríngeas | Ausente | Ausente | Presente | Ausente | Presente | Ausente |
*Algunas estructuras son metaméricas en ciertos moluscos primitivos, como Neopilina.
**Excepto en Urocordados.
1.- ¿Qué sietes caracteres pueden ser considerados como sinapomorfías? (recuerda, deben ser derivados y compartidos)
1.-
2.-
3.-
4.-
5.-
6.-
7.-
2.- A partir de la tabla de sinapomorfías, ¿cuál de estos cladogramas resulta más parsimonioso?
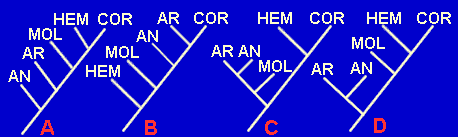
3.- ¿Existe algún caso de homoplasia en dicho cladograma? ¿cuál?
4.- ¿Cuál es la longitud del árbol B? ¿Y la del árbol C?
5.- ¿Cuál es el Indice de consistencia del cladograma de mayor parsimonia?
Tema 5.- Cladogramas y clasificación.
En este tema veremos cómo es posible aplicar el método filogenético al sistema linneano, un sistema que fue diseñado en un contexto preevolutivo, un contexto muy diferente al que estamos describiendo aquí.
Antes de desarrollar el tema, es preciso comprobar que habéis comprendido una propiedad importante de la Sistemática Filogenética. Un mismo carácter puede ser sinapomorfía, simplesiomorfía y autopomorfía a diferentes niveles taxonómicos. ¿Cómo es esto posible? Vamos a verlo con un ejemplo.
Tomemos como ejemplo el carácter "presencia de vértebras". Este carácter no es apomorfo ni plesiomorfo en sí mismo, sino en relación a un contexto taxonómico determinado. Veamos tres situaciones:
1.- Estamos trabajando sobre las relaciones filogenéticas de los Deuteróstomos, es decir, Equinodermos, Hemicordados, Urocordados, Cefalocordados y Vertebrados. Es obvio que la presencia de vértebras será un carácter derivado y no compartido (sólo tienen vértebras los vertebrados). Por tanto será una autopomorfía.
2.- Trabajamos ahora con las relaciones filogenéticas de los siguientes taxones: Urocordados, Cefalocordados, Elasmobranquios, Actinopterigios, Tetrápodos. La presencia de vértebras se da en tres de estos taxones (Elasmobranquios, Actinopterigios, Tetrápodos). Por tanto es un carácter derivado y compartido (una sinapomorfía).
3.- Ahora estamos a otro nivel, examinando las relaciones filogenéticas entre Lisanfibios, Lepidosaurios, Cocodrilos, Aves y Mamíferos, con los Elasmobranquios como grupo externo. Las vértebras han pasado a ser un carácter primitivo y compartido por todos los taxones, es decir, una simplesiomorfía.
Las tres posibilidades se reflejan en los siguientes cladogramas:
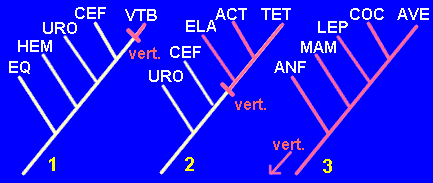
Como puede comprobarse, sólo en el caso 2, en el que la presencia de vértebras se convierte en una sinapomorfía, resulta informativa, es decir, útil para la construcción del cladograma.
Una vez que esto queda claro, podemos entrar en la cuestión de cómo debemos clasificar. La primera conclusión, que ya exponíamos en el tema 2, es que sólo los grupos estrictamente monofiléticos pueden ser considerados como taxones. Entonces definimos a los grupos monofiléticos como "todos y cada uno de los descendientes de un ancestral común". Ahora podemos añadir que el grupo monofilético es el definido por sinapomorfías, puesto que son las sinapomorfías las que nos hacen elaborar una hipótesis sobre un ancestral común a todos los miembros de un taxón. De esta forma hacemos que la definición descanse sobre nuestra herramienta de trabajo, la sinapomorfía.
¿Significa esto que los taxones, al igual que los cladogramas y las sinapomorfías, son provisionales? Evidentemente, esto es así. Un taxón, en Sistemática Filogenética, ímplica una hipotesis de relaciones filogenéticas y las hipótesis son, por definición, provisionales. Por esto, el panorama que nos dibuja la Sistemática Filogenética es el de una clasificación dinámica, sujeta a cambios pero, al mismo tiempo, organizada de acuerdo con criterios objetivos. Cualquier propuesta de clasificación debe justificarse, tanto con sinapomorfías como en lo que se refiere al criterio de máxima parsimonia.
| Peces |
| Anfibios |
| Reptiles |
| Aves |
| Mamíferos |
Todo esto ha cambiado radicalmente algunos esquemas tradicionales de clasificación. Recordemos la clasificación clásica de los vertebrados, de raíz linneana, que se muestra en esta tabla. Desde el punto de vista de la Sistemática Filogenética sólo las Aves y los Mamíferos serían taxones aceptables, en tanto en cuanto son monofiléticos. Una clasificación coherente con la Sistemática Filogenética requiere el establecimiento de taxones estrictamente monofiléticos, como ya veremos. Lo mismo sucede con ciertas agrupaciones a nivel suprafilético (por encima del phylum), tales como "acelomados", "celomados" o "pseudocelomados", agrupaciones difícilmente justificables con sinapomorfías.
Antes de exponer un ejemplo de clasificación filogenética, es preciso discutir un punto que hemos pasado por alto. La Sistemática Filogenética proporciona un método de agrupación, pero en la clasificación no sólo consiste en agrupar sino también en asignar al grupo una determinada categoría taxonómica (familia, orden, subclase, etc.).
Es preciso reconocer que la Sistemática Filogenética no proporciona un criterio universal de asignación de categorías. La respuesta consiste en que lo importante es que la clasificación 1) se base en grupos monofiléticos, 2) refleje el parentesco filogenético en la medida de lo posible. Asignar a un grupo la categoría de superorden o subclase se convierte así en mucho menos relevante que establecer las relaciones filogenéticas de dicho grupo con grupos cercanos. No se nos escapa que dicha asignación sigue conteniendo un componente subjetivo muy grande. Podemos prever que el cladograma, por tanto, irá ganando importancia en los libros de texto y artículos sobre Sistemática, en detrimento de los listados "lineales", cuyo contenido informativo es mucho menor.
Veamos qué taxones podemos distinguir en los vertebrados vivientes de acuerdo con los criterios de la Sistemática Filogenética. Un cladograma posible (provisional y sujeto a revisión, como todos los cladogramas) sería el siguiente:

*Lisanfibios: anfibios actuales. Existen muchas incertidumbres en lo que se refiere a la posición de holocéfalos, dipnoos, celacantos y quelonios.
Sólo se han indicado los principales taxones, mejor sustentados por sinapomorfías. Dichas sinapomorfías son las siguientes:
| Taxón: | Sinapomorfías: |
| Mandibulados | Presencia de mandíbulas |
| Teleóstomos | Mandíbulas secundarias (formadas por huesos dérmicos, no branquiales) |
| Tetrápodos | Miembro quiridio. Cintura pélvica fusionada a la columna vertebral |
| Amniotas | Amnios y alantoides |
| Diápsidos | Dos pares de aberturas temporales en el cráneo |
| Arcosaurios | Pelvis modificada. Ventana preorbitaria |
Los clásicos taxones (peces, anfibios, reptiles) quedan así sustituidos por los nuevos. Es sin duda una revolución, pero ¿acaso no fue una revolución la obra de Linneo?
Todo lo que hemos dicho hasta ahora tiene una importante consecuencia cuando examinamos el problema de la especie. En la Sistemática Evolutiva, el concepto de especie era muy importante. Ya expusimos en el tema 1 que la Sistemática Evolutiva propugnaba la utilización del concepto biológico de especie en lugar del concepto tipológico típico del sistema linneano.
La importancia de establecer un criterio a la hora de definir la especie se debe a los procedimientos usados tanto en el esencialismo linneano como en Sistemática Evolutiva. En ambos casos se trata de utilizar a la especie como entidad objetiva (realmente existente en la naturaleza) y unidad de clasificación. Una vez delimitada la especie, el procedimiento consiste en agrupar especies en taxones de categoría cada vez mayor (Géneros, familias, órdenes, etc.).
En cambio, dado que la Sistemática Filogenética parte del establecimiento de hipótesis sobre las relaciones filogenéticas, y no de la delimitación de especies, parece claro que el debate sobre el concepto de especie no es fundamental en Sistemática Filogenética, o al menos no es tan fundamental como en otras escuelas taxonómicas. Un ejemplo nos ayudará a comprender este punto.
Recordemos que la Sistemática Filogenética nos ayuda a establecer hipótesis de parentesco filogenético a cualquier nivel taxonómico, supra o infraespecífico. Supongamos que hemos estudiado la filogenia de cuatro poblaciones de animales (1,2,3 y 4) muy emparentadas entre sí, poblaciones que en términos tradicionales podrían ser consideradas especies próximas o "subespecies". Fruto de nuestro estudio podría ser el siguiente cladograma:
La figura de la izquierda representa el proceso real de filogenia de las cuatro poblaciones. Hemos señalado en rojo el momento en que la progresiva divergencia de los linajes origina la pérdida de la interfecundidad, es decir, individuos de las poblaciones 1 y 2 son interfecundos, pero los individuos de las poblaciones 3 y 4 sólo pueden reproducirse con los de la misma población.
Si adoptamos el criterio de especie biológica, parece claro que estamos ante tres especies (1+2, 3 y 4). Si dicho criterio no fuera aplicable por cualquier razón, es posible que una cierta diferenciación morfológica pudiera llevar a interpretar las poblaciones 1 y 2 como dos especies diferentes. Queda claro, por tanto, que la elección del criterio de especie puede suponer cambios en la clasificación. Sin embargo, ¿la elección de un criterio de especie u otro supondría cambios en el cladograma de la derecha? Parece obvio que no.
Recuerda: La elección de un determinado criterio de especie no influye en el establecimiento de relaciones de parentesco.
No quiere esto decir que la delimitación de especies no sea importante, ni mucho menos. Para el biólogo es imprescindible saber a qué especie pertenece el organismo con el que está trabajando. Lo que queremos decir es que dicha tarea de delimitación específica se ha convertido en una empresa relativamente independiente de la determinación de relaciones de parentesco, relaciones que son las que finalmente van a nutrir la clasificación de los seres vivos.
Ejercicios del tema 5
Este es un posible cladograma de los Deuteróstomos, los organismos con boca no derivada del blastoporo. A partir de este cladograma, responde a las siguientes cuestiones:

1.- ¿Cuáles de estos agrupamientos pueden constituir taxones en Sistemática Filogenética, cuáles no y por qué no?
| Hemicordados (Enteropneustos+Pterobranquios) | |
| Cordados (Urocordados+Cefalocordados+Vertebrados) | |
| Procordados (Urocordados+Cefalocordados) | |
| Cyrtotreta (Equinodermos+Enteropneustos+Pterobranquios+ Urocordados+Cefalocordados+Vertebrados) |
Taxones válidos:
Taxones no válidos:
No son válidos porque:
2.- De acuerdo con el cladograma, la presencia de notocorda sería una...(sinapomorfía, simpleisomorfía o autopomorfía)
Para determinados grupos de Deuteróstomos que figuran en el cladograma.
-
Para los diferentes grupos de Vertebrados.
-
Si consideramos en el cladograma a los Cordados como taxón terminal (formando una única rama).
-
3.- Algunos autores consideran a los Lofoforados (Foronídeos, Braquiópodos y Ectoproctos) como un grupo monofilético definido prácticamente por una sola sinapomorfía: el lofóforo, una corona de tentáculos peribucales con expansiones del mesocele. Sin embargo, los Pterobranquios tienen una estructura que responde a estas mismas características. Si suponemos que se trata de una estructura homóloga, no derivada de una convergencia (aparición independiente), como parece probable:
A) ¿Cómo podemos explicar la presencia de lofóforo en los pterobranquios sin moverlos del lugar que ocupan en el cladograma?
B) ¿Qué consecuencias tendría dicha explicación para la clasificación de los Deuteróstomos?
Tema 6.- Aplicaciones de la Sistemática Filogenética. Introducción a la Filogenia Molecular.
La filogenia molecular consiste en la obtención de información filogenética a partir del estudio de secuencias moleculares, es decir, ácidos nucleicos (DNA, RNA) o proteínas. Vamos a mostrar aquí cómo los métodos de la Sistemática Filogenética tienen una gran aplicación en filogenia molecular. Muchos piensan que la base de la filogenia molecular consiste en la comparación directa de secuencias, de forma que se consideran más relacionados los organismos en la medida en que sus secuencias son similares. Esta idea (que no es totalmente cierta, como luego veremos) tiene sus raíces en dos argumentos:
1.- Intuitivo. Parece obvio que cuando dos linajes biológicos divergen y evolucionan independientemente sus secuencias macromoleculares acumulan diferencias progresivamente. A más divergencia, más diferencia.
2.- Histórico. De hecho en el pasado se aplicaron técnicas que cuantificaban el grado de homología entre dos secuencias, y utilizaban dicho índice para establecer relaciones de parentesco entre organismos.
De nuevo nos encontramos en este campo el mismo dilema que el que exponíamos en el tema 1 sobre los fundamentos de la Sistemática, es decir, similitud versus parentesco filogenético. ¿Debemos considerar como medida de la divergencia filogenética entre dos organismos el grado absoluto de similitud entre las secuencias o debemos hacer un "análisis de caracteres" como el que hemos realizado en anteriores temas? ¿Podemos distinguir sinapomorfías en los caracteres moleculares? Vamos a aclarar este punto con un ejemplo.
Para simplificar, vamos a considerar el caso de tres organismos en los que compararemos una pequeña secuencia (homóloga, evidentemente) de cuatro bases:
| Organismo | Secuencia |
| 1 | AGCC |
| 2 | AGCT |
| 3 | GGTT |
Para trabajar con secuencias es frecuente comenzar con un procedimiento que no suele aplicarse en el caso de caracteres morfológicos, el de elaborar un árbol "no enraizado" (unrooted tree). La idea es construir un "árbol" conectando los taxones mediante ramas, pero de forma que las conexiones impliquen el menor número posible de cambios (otra vez el criterio de parsimonia). En el caso de nuestro ejemplo, tenemos que el árbol no enraizado de máxima parsimonia es:
Las barras verticales representan los pasos o cambios, tres en total. De momento no sabemos en qué dirección se producen dichos cambios, si la sustitución de la izquierda, en la cuarta posición de la secuencia, es en el sentido C->T o T->C. Lo que sí sabemos es que cualquier otra alternativa es menos parsimoniosa, por ejemplo:


Nos quedamos, por tanto con la primera opción. Pues bien, podemos avanzar más si somos capaces de "enraizar" al árbol, operación posible si conocemos la secuencia de bases en un grupo externo. Esto nos permitiría formular una hipótesis sobre la dirección de los cambios. Supongamos que en el grupo externo elegido la secuencia es AGCC. En este caso ya podemos construir un árbol enraizado que es, en realidad, un cladograma:
Ahora nos movemos ya por un terreno conocido. En efecto, la consideración de la secuencia AGCC como ancestral nos ha permitido identificar como derivados o apomorfos los cambios C->T en la cuarta posición, A->G en la primera posición y C->T en la tercera posición. Sin embargo, de estos tres cambios, sólo el primero, la sustitución de la citosina por la timina en la cuarta posición constituye un carácter derivado y compartido, es decir, una "sinapomorfía molecular" que nos permite sugerir una mayor relación de parentesco entre los organismos 2 y 3. En el fondo el procedimiento es el mismo que con los caracteres morfológicos, aunque la complejidad del análisis de las secuencias macromoleculares, con cientos o miles de posiciones, requiere el uso de ordenadores y algoritmos que funcionan de acuerdo con el razonamiento aquí expuesto. Los programas de análisis filogenético mediante máxima parsimonia buscan los árboles más cortos (con menos pasos) y efectúan un enraizamiento con respecto a la secuencia propuesta como ancestral.
Es muy importante que en nuestro ejemplo comparemos el resultado final, expresado en el cladograma, con el que habríamos obtenido a partir de la aplicación de un método de distancia (similitud). En efecto, el porcentaje de identidad entre las secuencias es el siguiente:
| AGCT | GGTT | |
| AGCC | 75%(3 de 4) | 25% (1 de 4) |
| GGTT | 50% (2 de 4) |
A la derecha tenemos un fenograma (ver tema 1) que representa la similitud (en tanto por uno) mediante el algoritmo UPGMA. Puede comprobarse como la menor distancia se da entre los organismos 1 y 2, que comparten el 75% de la secuencia.
Es decir, ¡los métodos de distancia hubieran dado un resultado completamente diferente al obtenido por el método de parsimonia! En efecto, un análisis basado en la similitud de las secuencias se hubiera centrado en los caracteres plesiomorfos (no informativos) A y C en la primera y tercera posición. Sin embargo, el único carácter con información filogenética, la T de la cuarta posición, hubiera pasado desapercibida.
Normalmente, el análisis de secuencias no será nunca tan sencillo como en el ejemplo anterior. La homoplasia, es decir, el número total de convergencias y reversiones, será importante, especialmente cuando trabajemos con secuencias de nucleótidos. En este caso, al manejar sólo cuatro letras, la probabilidad de que coincidan de forma independiente, o de que se retorne al estado ancestral, es relativamente alta. Por esto, en filogenia molecular trabajaremos frecuentemente con longitudes de árbol y con índices de consistencia, que nos darán la medida de la "bondad" de las agrupaciones.
Ejercicios del tema 6
Vamos a trabajar con una corta secuencia de cinco posiciones en cinco taxones diferentes. La tabla correspondiente es la siguiente:
| Taxón 1 | Taxón 2 | Taxón 3 | Taxón 4 | Taxón 5 |
| ACCCC | AGCCT | ACGCC | GGCCC | GGCAC |
1.- ¿Cuál de estos árboles no enraizados es más corto, es decir, implica menos número de cambios?(1, 2, 3 o 4)
2.- ¿Cuál es la longitud de cada árbol?
Arbol 1 : ____ pasos
Arbol 2 : ____ pasos
Arbol 3 : ____ pasos
Arbol 4 : ____ pasos
3.- Si el grupo externo tiene una secuencia AGCCT, ¿cuál de estos cladogramas será más parsimonioso?

4.- A la vista del cladograma que has elegido, ¿Cuáles de estos cambios contienen información filogenética? (Son "sinapomorfías moleculares").(si o no)
Adenina por guanina en posición 1 _____ contiene información.
Timina por citosina en posición 5 _____ contiene información.
Citosina por adenina en posición 4 _____ contiene información.
Guanina por citosina en posición 2 _____ contiene información.
Diccionario de términos
Ancestral común: Antepasado compartido por dos o más linajes o taxones. Dado que se admite un origen único de la vida, se sigue que todos los seres vivos sin excepción tienen algún ancestral común. Por este motivo se suele utilizar el concepto de "ancestral común más reciente" o "cenancestro", indicando el último ancestral común antes de la divergencia de los linajes. En Sistemática Filogenética los ancestrales comunes son hipotéticos, nunca se refieren a especies concretas.
Apomorfo: Se dice del estado de carácter avanzado o derivado en relación a otro primitivo. Véase sinapomorfía.
Autopomorfía. Carácter derivado, presente en un taxón concreto, pero no compartido. Por este motivo no contiene información filogenética.
Carácter: Atributo presente en un grupo de organismos, como consecuencia de la expresión de su genotipo durante el desarrollo. En Sistemática Filogenética se usan, en general, caracteres de tipo morfológico.
Categoría: Nivel ocupado por un taxón en la jerarquía linneana (p.e. especie, orden, subclase o phylum).
Cladograma: Hipótesis de relaciones de parentesco entre grupos monofiléticos. Adopta la forma de un árbol con ramificaciones dicotómicas o nodos. Al final de cada rama se sitúan los taxones terminales. Cada nodo debe estar justificado al menos con una sinapomorfía.
Consistencia: Medida de la bondad de un cladograma. Véase Indice de consistencia.
Convergencia: Véase Homoplasia.
Enraizamiento: Designación de determinados estados de carácter como ancestrales, con lo cual se asume una dirección para los cambios y queda "enraizado" el cladograma.
Esencialismo: Dejando de lado aspectos puramente filosóficos, el esencialismo en Sistemática propugna la definición de taxones a partir de caracteres "esenciales" o diagnósticos, lo que lleva a una clasificación basada en similitudes y diferencias, o a un concepto tipológico de especie. La Sistemática Filogenética no es esencialista, puesto que la clasificación se basa en relaciones de parentesco.
Especie biológica: Concepto de especie basado en una propiedad biológica como es la reproducción. En este contexto se concibe a la especie como una comunidad de individuos real o potencialmente interfecundos.
Especie nominalística: Determinados naturalistas (p.e. los racionalistas) negaron la existencia real de especies en la naturaleza, afirmando que cualquier agrupación de individuos con características similares resulta arbitraria. Negaban, por tanto, que hubiera discontinuidades reales, reproductivas o morfológicas entre dichas agrupaciones arbitrarias. Según este enfoque, la especie no sería más que un nombre (de ahí el término nominalismo) aplicado a un grupo de individuos para comodidad del taxónomo.
Especie tipológica: Concepto de especie basado en caracteres diagnósticos, "esenciales", fundamentalmente morfológicos, que distinguen a dicha especie de cualquier otra (véase esencialismo).
Estado de carácter: En Sistemática Filogenética suele hacerse la distinción entre el carácter y el estado del carácter, es decir, todas las distintas formas o variables que puede adoptar un atributo determinado. De esta forma el carácter es "número de dedos", y los estados del carácter son "0,1,2,3,4,5, etc.". En general, no es necesario hacer esta distinción y podemos hablar sólo de "caracteres".
Evolución: Disciplina que se ocupa de los procesos que generan cambios a lo largo de los linajes de los seres vivos. No confundir con Filogenia.
Feneticismo: Escuela sistemática que propugna la agrupación utilizando exclusivamente la similitud, generalmente mediante estimaciones cuantitativas de dicha similitud.
Fenograma: Representación gráfica de una semimatriz de índices de distancia en los procedimientos feneticistas. En el fenograma, los taxones se agrupan en función de su similitud global.
Filogenia: Denominamos así al patrón de relaciones de parentesco de los organismos o, dicho de otra forma, al estudio de las relaciones entre los linajes de los organismos bajo una perspectiva histórica.
Grupo hermano: Para un grupo dado G, el grupo hermano es el que parte del mismo nodo que G. Dicho de otra forma, dos ramas que parten de un mismo nodo constituyen grupos hermanos. En la figura el grupo hermano de B es A, y el de C es D, y viceversa. Además, el grupo hermano de C+D es A+B, y viceversa.
Grupo externo: Grupo de organismos no perteneciente al grupo que está siendo analizado. Los grupos externos proporcionan información acerca de los estados plesiomorfos (ancestrales) de los caracteres, puesto que los estados apomorfos (derivados) no deberían aparecer en grupos externos. Siempre es conveniente elegir grupos externos que estén tan emparentados con el que analizamos como sea posible.
Grupo monofilético: Véase monofilia.
Homoplasia: Similitud no debida a parentesco. La homoplasia puede deberse a la convergencia o aparición independiente de dos caracteres derivados (p.e. alas en aves y murciélagos), pero también a la reversión o pérdida de un carácter derivado (ausencia de patas en peces y cetáceos). En la construcción de cladogramas es posible admitir casos de homoplasia, pero es obligado, de acuerdo con el criterio de parsimonia, elegir el cladograma que minimice el número de dichos casos.
Indice de consistencia: Se define como el cociente entre el número de cambios presentes en la tabla de caracteres y el número de cambios que se producen a lo largo de un cladograma determinado. Si no hay homoplasia, dichos números serán iguales y el índice de consistencia será igual a 1. De acuerdo con el principio de parsimonia, debe elegirse siempre el cladograma con mayor índice de consistencia.
Inferencia filogenética: Proceso de razonamiento por el cual se elabora una hipótesis sobre las relaciones de parentesco entre los taxones incluidos en un grupo monofilético. El proceso incluye el análisis de caracteres, la determinación de los estados de carácter derivados y compartidos (sinapomorfías), y la construcción de un cladograma.
Longitud del árbol: Número de cambios en los caracteres que se producen a lo largo de las ramas de un cladograma. El principio de parsimonia obliga a la elección de los cladogramas de menor longitud, puesto que serán los que impliquen menor homoplasia.
Monofilia: En Sistemática Filogenética un grupo será monofilético si, y solo si, comprende a todos los taxones derivados de un único ancestral común. Dicho de otra forma, un grupo monofilético comprende a un ancestral común y todos sus descendientes. El criterio de monofilia estricta nos dice que todos los taxones deben ser estrictamente monofiléticos.
Nodo: Véase cladograma.
Parentesco filogenético: El grado de parentesco filogenético entre dos taxones determinados es proporcional a la distancia, en el tiempo, a su más reciente ancestral común. De esta forma dos taxones A y B están más emparentados entre sí que cualquiera de ellos con un tercero C, si A y B comparten un ancestral común más reciente que los ancestrales comunes más
recientes de A con C y de B con C. El parentesco filogenético es un criterio de agrupación en Sistemática.
Parsimonia: Principio metodológico que postula la elección de la hipótesis más sencilla entre varias hipótesis concurrentes. Aplicado a la Sistemática Filogenética el criterio de parsimonia conduce a la elección del cladograma que explica la filogenia de un grupo con el menor número de cambios, es decir, con la menor homoplasia posible.
Plesiomorfo: Se dice del estado de carácter primitivo o ancestral en relación a otro derivado. Véase simplesiomorfía.
Polaridad: Sentido del cambio, es decir, elaboración de una hipótesis acerca del orden relativo de aparición de los caracteres a lo largo de la filogenia del grupo. De esta forma postulamos que la polaridad del cambio pata/ala en los tetrápodos es en el sentido pata -> ala y no al revés. Esto convierte a la presencia de alas en un estado de carácter apomorfo (derivado) respecto a la presencia de patas.
Reversión: Véase Homoplasia.
Similitud: Utilizamos aquí este término en el sentido de la semejanza morfológica entre individuos o taxones. Constituye uno de los dos criterios de agrupación en Sistemática.
Simplesiomorfía: Carácter primitivo y compartido. Al ser primitivo, heredado de un ancestral lejano en el tiempo, no contiene información filogenética ni debe utilizarse para hacer agrupaciones.
Sinapomorfía: Carácter derivado y compartido por dos o más taxones. Las sinapomorfías definen grupos monofiléticos y son la principal herramienta de trabajo en Sistemática Filogenética. Cada nodo de un cladograma debe estar justificado por al menos una sinapomorfía.
Sistemática: En sentido estricto es sinónimo de taxonomía. En sentido amplio es la disciplina que se ocupa de la diversidad de los seres vivos.
Sistemática filogenética: Escuela sistemática, fundada por el entomólogo alemán Willi Hennig, que propugna la agrupación de taxones exclusivamente a partir del parentesco filogenético.
Taxón: Unidad de clasificación, formada por un grupo de organismos con características definitorias (p.e. Angiospermas, Gasterópodos, Rana perezi, homínidos...). En Sistemática Filogenética dichas características deben ser, obligatoriamente, sinapomorfías.
Taxonomía: Disciplina biológica que se ocupa de la clasificación de los seres vivos, es decir, de su nomenclatura y ordenamiento en taxones.
Taxonomía numérica: Véase feneticismo.
CRÉDITOS
La elaboración del curso ha sido posible gracias al proyecto de Innovación Educativa "Elaboración de material interactivo y multimedia para la enseñanza de la Sistemática Filogenética" concedido por el Instituto de Ciencias de la Educación de la Universidad de Málaga para el curso 1998-99.
MET. Y TEC. EXP. EN BIOLOGIA ANIMAL M.G.S. (2002)
50
Descargar
| Enviado por: | Canija |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.