Biología, Botánica, Genética y Zoología
Permeabilidad y transporte pasivo en membranas celulares
MEMBRANAS CELULARES: PERMEABILIDAD Y TRANSPORTE PASIVO
Abstract.
Sometimos diversos cilindros de remolacha (Beta vulgaris L.) a distintas condiciones con el fin de estudiar los efectos de varios tratamientos químicos y físicos sobre la permeabilidad de las membranas celulares.
Por otra parte utilizamos cilindros de patata (Solanum tuberosum L.) incubados previamente en una solución de sacarosa para mediante el método del Q10 demostrar que el transporte del agua a través de las membranas es de tipo pasivo debido al gradiente de potencial hídrico, es decir, un mecanismo osmótico.
Los resultados obtenidos permiten estimar un aumento en la permeabilidad de las membranas tras someterlas a varios tratamientos físicos y químicos.
Palabras clave: permeabilidad, transporte pasivo, Beta vulgaris, Solanum tuberosum, espectrofotómetro, método del Q10, absorbancia.
Introducción.
Composición de la membrana:
Entran a formar parte lípidos, proteínas y glúcidos en proporciones aproximadas de 40%, 50% y 10%, respectivamente.
Lípidos: en la membrana de la célula eucariota encontramos varios tipos de lípidos. Fosfolípidos, glucolípidos, sulfolípidos y colesterol. Todos tienen carácter anfipático ; es decir que tienen un doble comportamiento, una parte de la molécula es hidrófila y la otra es hidrófoba, por lo que cuando se encuentran en un medio acuoso se orientan formando una bicapa lipídica . La membrana plasmática no es una estructura estática, sus componentes tienen posibilidades de movimiento, lo que le proporciona una cierta fluidez. Los movimientos que pueden realizar los lípidos son rotación, difusión lateral, flip-flop y flexión.
La fluidez es una de las características más importantes de las membranas y depende de factores como: la temperatura, a medida que esta se incrementa, la fluidez aumenta. Y la naturaleza de los lípidos, la presencia de lípidos insaturados y de cadena corta favorecen el aumento de fluidez; la presencia de colesterol endurece las membranas, reduciendo su fluidez y permeabilidad.
Proteínas: son los componentes de la membrana que desempeñan las funciones específicas de transporte, comunicación, etc. Las proteínas de membrana se clasifican en: proteínas integrales, están unidas a los lípidos íntimamente, suelen atravesar la bicapa lípidica una o varias veces, por esta razón se les llama proteínas de transmembrana. Y proteínas periféricas, que se localizan a un lado u otro de la bicapa y están unidas débilmente a las cabezas polares de los lípidos de la membrana u a otras proteínas integrales por enlaces de hidrógeno.
Glúcidos: se sitúan en la superficie externa por lo que contribuyen a la asimetría de la membrana. Estos glúcidos son oligosacáridos unidos a los lípidos (glucolípidos), o a las proteínas (glucoproteínas). Esta cubierta de glúcidos constituye el glucocálix, el cual desempeña algunas funciones como la protección de la superficie de las células de posibles lesiones, conferir viscosidad a las superficies celulares, permitiendo el deslizamiento de células en movimiento, presentar propiedades inmunitarias, etc.
Estructura:
En la actualidad el modelo más aceptado para explicarla, es el propuesto por Singer y Nicholson, denominado modelo del mosaico fluido, que presenta las siguientes características: considera que la membrana es como un mosaico fluido en el que la bicapa lipídica es la red cementante y las proteínas están embebidas en ella, interaccionando unas con otras y con los lípidos. Tanto las proteínas como los lípidos pueden desplazarse lateralmente. Los lípidos y las proteínas integrales se hallan dispuestos en mosaico. Las membranas son estructuras asimétricas ya que en ambas partes hay una diferente distribución de los componentes; así, por ejemplo los glúcidos, sólo se encuentran en la cara externa (Alberts y col., 1992).
Permeabilidad y transporte:
Debido a su composición, la membrana plasmática es semipermeable y por lo tanto el transporte de sustancias a través de membrana depende de varios factores entre los que destacan la polaridad y el tamaño:
En cuanto a la polaridad, mientras las sustancias apolares atraviesan la membrana sin ningún problema, las moléculas polares necesitan ser transportadas para poder atravesar la parte hidrofóbica de la membrana debida a las colas de los ácidos grasos. También decir que el agua, pese a que es una molécula polar, es la que más fácilmente atraviesa la membrana, gracias a unos poros denominados acuoporinas
Y respecto al tamaño, un elevado peso molecular implica que las moléculas no puedan atravesar la membrana.
Por tanto, a través de estas membranas se realiza el transporte de las sustancias necesarias para el metabolismo celular. Este transporte puede ser activo o pasivo, en función de que se lleve a cabo en contra o a favor, respectivamente, del gradiente electroquímico, lo cual va asociado a la utilización directa o no de energía metabólica.
Transporte activo:
La célula utiliza ATP como fuente de energía para ayudar a las moléculas a atravesar la membrana, a través de proteínas translocadoras.
El transporte activo puede dividirse en:1) primario, cuando el transporte está ligado directamente a una reacción química como la hidrólisis de ATP.2) secundario, donde la translocación de la especie en contra del gradiente electroquímico está acoplada al transporte de otra especie a favor de dicho gradiente, de modo que la magnitud absoluta de la energía libre es lo suficientemente alta para impulsar el transporte de ambas.
El transporte mediado también puede clasificarse según la estequiometría del proceso en: uniporte cuando sólo se transporta una sola molécula, simporte cuando se transportan dos moléculas a la vez en el mismo sentido, y antiporte cuando se transportan dos moléculas en sentido opuesto. (Azcón y Talón., 2001).
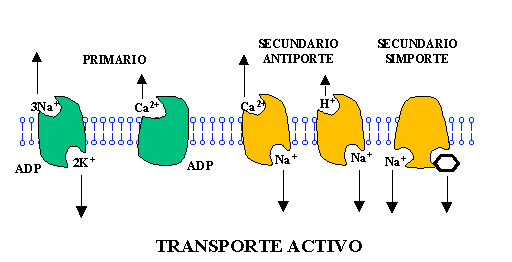
Fig 2. Mecanismos de transporte activo.
Transporte pasivo:
No se requiere energía para que la sustancia cruce la membrana plasmática. Los mecanismos de transporte pasivo son: difusión simple, osmosis, ultrafiltración , difusión facilitada.
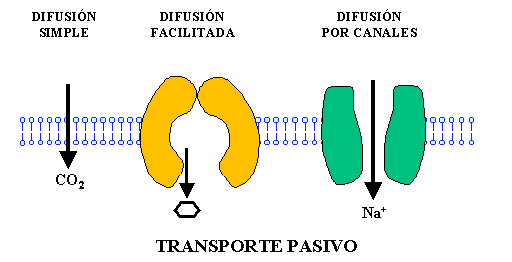
Fig. 3. Mecanismos de transporte pasivo.
Cuando hablamos de transporte pasivo debemos separar las partículas de intercambio entre nutrientes y agua, pues son procesos diferentes pero complementarios.
En el transporte pasivo de nutrientes se denomina difusión simple y cumple con las siguientes características: 1.el transporte se realiza sin gasto de energía, 2. el transporte es a favor de la gradiente de concentraciones de soluto; siempre desde un lugar con mayor concentración a un lugar de menor concentración, 3.el transporte se realiza de un medio hipertónico a un medio hipotónico.
La difusión simple se puede realizar directamente con la doble capa de fosfolípidos o a través de proteínas transmembranas. Un ejemplo es el que los lípidos y solventes orgánicos (alcohol) pasan a través de la capa de lípidos, por el fenómeno de afinidad. Normalmente un solvente orgánico atraviesa las membranas con facilidad. Las demás moléculas e iones atraviesan la membrana por diferencia de tamaño; las de menor peso molecular atraviesan las membranas por los canales con relativa facilidad
El transporte pasivo de agua se denomina osmosis y ella involucra al principal solvente inorgánico, el H20; implica la movilización del H20. El proceso de osmosis cumple con las mismas propiedades que la difusión simple.
Varios autores han utilizado el método de la Q10 para determinar el tipo de transporte que se produce en las membranas celulares, no sólo a nivel vegetal. Como por ejemplo, para determinar el tipo de transporte del zinc en un biota acuático viendo así su dependencia con la temperatura ( Glover y col., 2003). También se ha experimentado con la raiz del arroz comprobando con este método que utilizan un transporte pasivo para el ion amonio (Wang y col., 1993).
No mediado
Pasivo (canales y portadores)
TRANSPORTE
Termodinámica
Mediado Primario (bombas)
Activo
Secundario (portadores)
Uniporte
Estequeometría Simporte
Antiporte
Fig. 4. Esquema tipos de transporte.
En el transporte pasivo la velocidad se incrementa ligeramente cuando se eleva la temperatura 10º C. Pero el transporte pasivo un incremento de 10º C aumenta el doble o más la velocidad, hasta una cierta temperatura, ya que la mayoría de los enzimas se desnaturalizan a 50º C.
Valores de Q10 entre 1.2 y 1.5: mecanismos de transporte pasivo.
Valores de Q10 entre 2 y 4: mecanismos de transporte activo.
Valores de Q10 mayores de 4: desnaturalización de proteínas
Para llevar a cabo esta práctica utilizamos raíz de remolacha (Beta vulgaris L). Esta planta está compuesta por sacarosa (15-20%), pigmentos, fundamentalmente betacianina (rojo-púrpura), colina, glutamina, vitaminas A, B, C, sales minerales y fitoestrógenos. Las células contienen grandes cantidades de betacianina en sus vacuolas.
En condiciones normales el tonoplasto y el plasmalema son impermeables a este pigmento pero si aplicamos los tratamientos citados anteriormente podemos aumentar su permeabilidad y así permitir la salida del pigmento de las células. Después de este proceso medimos la absorbancia en el espectrofotómetro para así cuantificara el aumento de permeabilidad.
Existen otros factores que también dañan las membranas, entre ellos:
Los detergentes: son los agentes más útiles que alteran las propiedades bioquímicas de la membrana. Forman pequeñas moléculas anfipáticas que tienden a formar micelas en el agua. Al mezclarlos con las membranas, los extremos hidrofóbicos de las moléculas de detergente, se unen a las regiones hidrofóbicas de la zona externa de las proteínas de membrana, desplazando así las moléculas lipídicas. Puesto que el otro extremo de la molécula de detergente es polar, la unión detergente-proteína tiende a disolver en agua a las proteínas de membrana.
En resumen, las proteínas transmembrana y también otras proteínas estrechamente unidas a la membrana, pueden ser solubilizadas por estos detergentes destruyendo la bicapa lipídica (Albert y col., 1992).
Algunos ejemplos de detergentes son: Dodecil sulfato sódico (SDS), detergentes del tipo Tritón, Tween 20.
Cambios en el pH: realizan la misma función que los disolventes orgánicos; interrumpiendo las proteínas en la membrana, lo que causa que el contenido se difunda a la solución externa (White.,2004).
Material y métodos.
Para la determinación de la permeabilidad de membrana y el tipo de transporte dividimos la práctica en dos apartados bien diferenciados:
Permeabilidad de membrana:
En esta primera parte estudiamos de los efectos de los tratamientos físicos como la congelación y temperaturas altas (50º y 70º) y los tratamientos químicos como metanol y acetona sobre la permeabilidad de las membranas.
Para poder realizar la práctica primero cortamos con el sacabocados 6 cilindros de remolacha (sin corteza) de 1 centímetro de longitud y 1 centímetro de diámetro, que utilizaremos para medir el efecto de la temperatura sobre sus membranas. Para ello después de haberlos lacado con agua en durante 10-15 minutos, introducimos dos de los 6 cilindros en dos tubos atemperados a 70 y 50º C. Para después de 1 minuto trasladar los 2 cilindros a otros dos tubos con agua destilada (preparados previamente) junto con otro tubo con agua destilada y cilindro que nos servirá de control.
En el segundo de los tratamientos, después de descongelar un cilindro de remolacha colocarlo en un tubo de ensayo con 10 mililitros de agua destilada. Al cabo de 30-45 minutos determinar su absorbancia a 479 nanómetros.
Para saber el efecto de sustancias químicas sobre la permeabilidad de membrana preparamos 2 tubos de ensayo con 5 mililitros de metanol y 5 mililitros de acetona en cada uno de los cuales introdujimos un cilindro de remolacha durante 1 minuto, luego los trasladamos a 2 tubos con 10 mililitros de agua destilada. Transcurrido 45 minutos mediremos su absorbancia también a 479 nanómetros.
Determinación del Q10:
En la segunda parte comprobamos la existencia de transporte pasivo mediante el método del Q10. Para ello utilizaremos cilindros de patata (Solanum tuberosum L.) incubados previamente 24 horas en una solución de sacarosa 0.4 M.
En esta segunda parte comprobaremos si el transporte es pasivo utilizando el método del Q10. El cual hayamos mediante la formula:
(P4 - P3) / P3
Q10=
(P2 - P1) / P1
Para adquirir estos parámetros debemos coger 50 discos de patata incubados previamente 24 horas en una solución de sacarosa 0.4 M, para después secarlos ligeramente y pesarlos, esto será el valor de P1. A continuación separarlos en 2 grupos de 25 discos.
Primero secamos y pesamos los discos del grupo a 25º C (P1), luego los introducimos en un baño con agua destilada a 25º C durante 30 minutos, después de los cuales lo volvimos a secar y pesar (P2). Repetimos el proceso pero ahora secando y pesando los cilindros del grupo a 35º C (P3), previamente a introducirlos en un baño con agua destilada a 35º C y después de 30 minutos los volvimos a secar y pesar (P4).
Loe experimentos fueron realizados por 3 grupos independientes con 7-8 repeticiones en cada uno.
Resultados y discusión
Para analizar los resultados vamos a diferenciar entre los dos apartados de que constaba la práctica: los efectos de distintos tratamientos físicos (congelación y altas temperaturas) y químicos (metanol y acetona). Y la comprobación por el método Q10 que el transporte de las membranas es de tipo pasivo.
Permeabilidad de membranas:
Sabemos que la absorbancia es directamente proporcional a la concentración de pigmento, luego a mayor absorbancia, mayor rotura de las membranas.
Según los datos obtenidos por la media de los tres grupos, se observa que la congelación es el proceso que mas ha dañado las membranas celulares, ya que es el mayor valor de la absorbancia a 470 nm. Obteniendo una media de 1,75 ± 0,83 nm.
El efecto que hace la congelación se explica por la formación de cristales de hielo que rompen fácilmente las membranas. Estos cristales tan sólo necesitan una temperatura de -2Cº a -3Cº para formarse, y nuestra temperatura era de -20Cº, con lo cual la formación de los cristales es evidente (Azcón y Talón., 2001).
A continuación, el segundo proceso que más actúa en los tres grupos es el tratamiento con metanol, que es un proceso químico. El metanol es un disolvente orgánico, al igual que la acetona, y actúa rompiendo la estructura puesto que la membrana contiene un 30% de fosfolípidos solubles en etanol. Obteniéndose un valor medio de 0,52 ±0,29 nm.
Podemos observar una diferencia de absorbancia entre las membranas tratadas con metanol y las tratadas con acetona (los dos procesos químicos) del orden de 0,5nm.
En estos tratamientos debería actuar con mayor eficacia la acetona ya que es más apolar debido al doble enlace, por lo que disolvería más los lípidos, pero esta diferencia se puede explicar porque la acetona está más diluida.
Con respecto al efecto provocado por el resto de temperaturas, debemos señalar antes que tanto el frío como el calor provocan pérdida de semipermeabilidad ya que modifican la microviscosidad o fluidez de las membranas, el calor la reduce mientras que el frío la aumenta (Azón y Talón., 2001).
Esto afecta también al funcionamiento de las proteínas integrales de membrana porque dependen de que la membrana mantenga su fluidez.
En nuestros datos observamos que en todos los casos la absorbancia, da menores valores en membranas tratadas a altas temperaturas.
Si nos fijamos en los datos veremos que siempre da mayor absorbancia con membranas tratadas a 70Cº que las tratadas a 50Cº (con valores de más del doble), esto ocurre porque cuanto más alta sea la temperatura, más disminuirá la fluidez de la membrana lo que conlleva menos resistencia y más rotura.
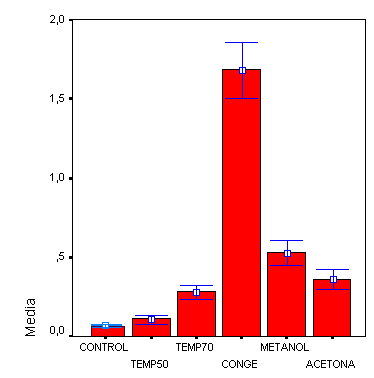
Histograma general
Estadísticos descriptivos generales
|
| N | Mínimo | Máximo | Media | Desv. típ. |
| CONTROL | 24 | ,035 | ,160 | ,07042 | ,031757 |
| TEMP50 | 23 | ,030 | ,470 | ,11204 | ,102970 |
| TEMP70 | 23 | ,020 | ,600 | ,28117 | ,167067 |
| CONGE | 24 | ,480 | 3,600 | 1,67875 | ,625073 |
| METANOL | 24 | ,150 | 1,500 | ,51792 | ,285459 |
| ACETONA | 24 | ,080 | ,950 | ,35467 | ,229213 |
| N válido (según lista) | 16 |
|
|
|
|
En general y observando el histograma de las medias de los tres grupos conjuntamente, vemos que el proceso que más ha dañado a las membranas es la congelación, seguido de los procesos químicos con etanol y acetona, y finalmente las altas temperaturas.
Determinación del Q10:
Observando los resultados de las tres medias para el Q10 de los tres grupos, no descartamos ningún dato, ya que todos se dieron por válidos en sus correspondientes prácticas, es decir; en todos los grupos se comprobó que efectivamente, había transporte pasivo.
Para reafirmarnos, realizamos la media de las tres Q10 obteniendo un valor de 1,14 ± 0,28 nm.
Los errores de experimentación se deben a factores externos, como las medidas de concentración, las diferentes características de los trozos de remolacha y patata, etc.
Pero nosotros no los hemos eliminado ya que no suponían cambios en el resultado
Bibliografía:
& AZCON-BIETO J., TALON, M. 2001. Fundamentos de fisiologia vegetal. Ed McGraw-Hill Interamericana. Madrid.
& ALBERTS B., BRAY D., LEWIS J., RAFF M., ROBERTS K., WATSON J.D. 1992. Biología molecular de la célula. Ed. Omega, Barcelona.
& WHITE I. 2004. Effect of salt and pH on the permeability of beet cells.
http:// www.madsci.org/posts/archives/2004-03/1079632628.Cb.r.html. Revista Madsci.
& NAVARRI-IZZO F. 2000.Protein dynamics in thylakoids of the desiccation tolerant plant Boea hygroscopica during dehydration and rehydration.
http:// www.plantphysiol.org/cgi/content/full/124/3/1427
& WANG MY., SIDDIQI MY., RUTH TJ., GLASS ADM. 1993. Ammonium uptake by rice root.2. kinetics of NH4+-N-13 influx across the plasmalemma. http://plantphysiology.com
& GLOVER CM., BURY NR., HOGSTRAND C. 2003. Zinc uptake across the apical membrane of freshwater rainbow trout intestine is mediated by high affinity, low affinity, and histidine-facilitated pathways. http://wos02.isiknowledge.com.
& AGUILAR M. Y CLAROS MG. Transporte de aminoácidos en vegetales. Departamento de bioquímica y biología molecular. Universidad de Málaga.
http://www.encuentros.uma.es/encuentros37/tranporte.html
ANEXO
| BL3 | Media |
| Control | 0,07 |
| 50º C | 0,10 |
| 70º C | 0,24 |
| Congelación | 1,74 |
| Metanol | 0,52 |
| Acetona | 0,41 |
| Q10 | 1,31 |
| BI1 | Media |
| Control | 0,05 |
| 50º C | 0,07 |
| 70º C | 0,29 |
| Congelación | 1,76 |
| Metanol | 0,46 |
| Acetona | 0,23 |
| Q10 | 1,12 |
| BI2 | Media |
| Control | 0,09 |
| 50º C | 0,16 |
| 70º C | 0,30 |
| Congelación | 1,53 |
| Metanol | 0,56 |
| Acetona | 0,41 |
| Q10 | 0,99 |
Histograma grupo 1.
Estadísticos descriptivos grupo1
|
| N | Mínimo | Máximo | Media | Desv. típ. |
| CONTROL | 8 | ,040 | ,070 | ,05437 | ,009797 |
| TEMP50 | 8 | ,035 | ,090 | ,07025 | ,019062 |
| TEMP70 | 8 | ,180 | ,400 | ,29125 | ,096427 |
| CONGE | 8 | 1,100 | 2,200 | 1,76250 | ,396187 |
| METANOL | 8 | ,240 | ,850 | ,46125 | ,201880 |
| ACETONA | 8 | ,080 | ,420 | ,23313 | ,130683 |
| Q10 | 5 | ,843 | 1,320 | 1,11860 | ,186560 |
| N válido (según lista) | 5 |
|
|
|
|
Histograma grupo 2
Estadísticos descriptivos grupo 2
|
| N | Mínimo | Máximo | Media | Desv. típ. |
| CONTROL | 8 | ,050 | ,140 | ,08688 | ,029147 |
| TEMP50 | 8 | ,040 | ,470 | ,16250 | ,135066 |
| TEMP70 | 8 | ,085 | ,510 | ,30313 | ,160066 |
| CONGE | 8 | ,480 | 2,300 | 1,52875 | ,630905 |
| METANOL | 8 | ,350 | ,800 | ,56375 | ,139482 |
| ACETONA | 8 | ,270 | ,580 | ,41188 | ,104879 |
| Q10 | 6 | ,821 | 1,290 | ,99483 | ,198441 |
| N válido (según lista) | 6 |
|
|
|
|
Histograma grupo 3
Estadísticos descriptivos grupo 3
|
| N | Mínimo | Máximo | Media | Desv. típ. |
| CONTROL | 8 | ,035 | ,160 | ,07000 | ,042003 |
| TEMP50 | 7 | ,030 | ,330 | ,10214 | ,106453 |
| TEMP70 | 7 | ,020 | ,600 | ,24457 | ,243436 |
| CONGE | 8 | ,820 | 3,600 | 1,74500 | ,830680 |
| METANOL | 8 | ,150 | 1,500 | ,52875 | ,448695 |
| ACETONA | 8 | ,090 | ,950 | ,41900 | ,345249 |
| Q10 | 6 | ,920 | 1,808 | 1,31050 | ,350444 |
| N válido (según lista) | 5 |
|
|
|
|
Descargar
| Enviado por: | Xicolisto |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.