Geología
Paleontología de invertebrados
Tema 2: Moluscos
Caracteres generales
Los moluscos son uno de los grupos fósiles más importantes de los invertebrados, tanto por su variabilidad morfológica como por la ecológica.
En general, son formas desprovistas de segmentación, con la excepción del grupo más antiguo, los monoplacóforos, que tienen un cuerpo fuertemente segmentado. Son metazoos, triblásticos, celomados y protostomios.
Están provistos originalmente de simetría bilateral, aunque en algunos grupos se pierde por procesos de torsión en la concha debido a adaptaciones secundarias.
El cuerpo de los moluscos se divide en cuatro partes:
-
Cabeza
Perfectamente desarrollada, posee casi siempre ojos y tentáculos.
-
Pie
En forma de suela, se sitúa en posición ventral. Función locomotora.
-
Masa visceral
Se sitúa en posición dorsal y en ella se incluyen todos los órganos fundamentales del molusco.
-
Manto
Es un extenso envoltorio tegumentario responsable de la formación de la concha.
-
Concha
La concha es de CaCO3 y a veces posee también entremezclado un material proteínico, la conquiolina. Está constituída por una sola pieza, por dos, o por ocho en el caso de los poliplacóforos, en cuyo caso están ligeramente articuladas. Este esqueleto pueda ser externo, interno, o no poseerlo.
La microestructura es fibrosa o prismática y la composición de la concha determina tres capas que desde el exterior hacia el interior se denominan:
-
Periostraco:
Capa curticular constituida por material proteínico exclusivamente que normalmente no llega a fosilizar.
-
Ostraco:
Es una capa de calcita o aragonito, o bien una mezcla de ambas junto con capas de material proteínico de tipo conquiolínico. Microestructura fibrosa o prismática.
-
Hipostraco:
Constituído por capas alternantes de CaCO3 y de conquiolina, también de naturaleza fibrosa y dispuestas formando un ángulo muy agudo con las capas del ostraco.
Clasificación
Es prácticamente coincidente en paleontología y en neontología, porque casi todos los grupos llegan hasta la actualidad. Los moluscos se dividen en las siguientes clases:
-
Monoplacóforos (segmentados)
-
Anfineuros
-
Estenotecoides
-
Escafópodos
-
Gasterópodos
-
Lamelibranquios o bivalvos
-
Cefalópodos
-
Tentaculítidos
Los moluscos han aparecido a partir del Cámbrico inferior, pero ya en este momento formas correspondientes a gasterópodos y monoplacóforos estaban relativamente evolucionadas, con lo que es de suponer un origen anterior (Precámbrico).
Monoplacóforos
Caracteres generales
Lo monoplacóforos son segmentados, lo cual indica que su origen ha de estar en un grupo de organismos segmentados, posiblemente los anélidos (tubelarios). Este grupo, junto con los anfineuros y los estenotecoides está poco representado en la columna, pero sus apariciones se relacionan con medios muy concretos.
Representados en la columna sólo desde el Cámbrico al Devónico, se creía que sólo existían fósiles pero hace algún tiempo aparecieron ejemplares similares a estos. Conchas univalvas, y simetría bilateral que sólo se observa en el interior se las conchas por las huellas musculares es lo más destacado.
Desde el punto de vista filogenético son importantes debido a que son los únicos que tienen un cuerpo segmentado.
Estenotecoides
Caracteres generales
Sólo representados por tres géneros, se creía que eran monoplacóforos, pero las conchas eran bivalvas y no parecía que estuviesen segmentados. Poca importancia desde el punto de vista paleontológico.
Anfineuros
Caracteres generales
Divididos en aplacóforos (de lo que no existen fósiles), y poliplacóforos, registrados a partir del Cámbrico superior. Son los únicos con esqueleto de ocho piezas, su cuerpo está alargado longitudinalmente y aplanado ventralmente. Aparecen en los sedimentos como placas diseminadas.
Actualmente están representados por un único grupo, los chitones, que aparecen en los acantilados. Se desarrollan en medios muy restringidos, son poco frecuentes en la columna estratigráfica y son poco importantes paleontológicamente.
Escafópodos
Caracteres generales
Representados por formas marinas bentónicas, están provistos de un esqueleto de CaCO3, de aspecto cónico, alargado y ligeramente curvado, abierto por la región anterior en una amplia apertura y en la posterior por una apertura mucho más fina, llamada escotadura.
Simétricos bilateralmente, la ornamentación se basa en estrías o costillas longitudinales y en ocasiones, anillos más o menos concéntricos relacionados con fases de crecimiento. Más raramente son lisos.
Actualmente, este grupo está representado por formas que viven semienterradas por su parte anterior en el barro de medios neríticos y poco profundos, próximos a la costa. Se cree que en el pasado debió de ser más o menos similar. Aparecen desde el Ordovícico hasta la actualidad, donde parece que se encuentran en plena expansión. Se consideran organismos excavadores.
Lamelibranquios o bivalvos
Caracteres generales
Son moluscos acéfalos, marinos y en algunas ocasiones, de agua dulce. Están adaptados a diferentes medios de vida:
-
Vida libre
Algunos tienen vida libre y se desplazan reptando mediante el pie o por propulsión, abriendo y cerrando violentamente las valvas.
-
Vida sedimentaria
Se fijan periódicamente al sustrato o a la concha de otros animales por medio de unos pelillos segregados por el biso o bien cementando una de las valvas.
-
Vida enterrada
Viven enterrados en la arena, fango o entre las rocas durante toda su vida.
Tienen el cuerpo protegido por una concha calcárea, bivalva y provista de simetría bilateral con respecto al plano de comisura, dando lugar a conchas equivalvas. Sólo en algunas ocasiones y por adaptaciones secundarias una de las valvas se deforma perdiendo así la simetría bilateral, dando lugar a conchas inequivalvas.
La orientación de la concha es tal que el borde articulante es considerado como región dorsal mientras que la parte opuesta es región ventral.
A lo largo de la región dorsal aparece un músculo ligamentario, que mantiene siempre las valvas abiertas, mientras que los músculos aductores hacen que la concha se cierre herméticamente. Aparte de estos músculos existen otros como los braquiales, relacionados con la inserción de las branquias, los bisales, con el biso y la inserción del manto, que deja lo que se llama en fósiles la línea paleal, los paleales.
En el manto también existen invaginaciones que se corresponden con la salida al exterior de un por de sifones, considerados como órganos carnosos cuya función es la de impeler o expeler corrientes de agua y otros productos hacia la cavidad del manto o paleal.
Los parámetros más representativos de la morfología de la concha son:
-
Longitud
Distancia entre dos planos perpendiculares al eje cardinal y que tocan las partes más salientes de la región anterior y posterior de la concha.
-
Altura
Distancia entre dos planos perpendiculares al plano comisural y que tocan las partes más salientes de las regiones dorsal y ventral.
-
Inflación
Distancia que existe entre dos planos paralelos al plano de comisura y que tocan las partes más convexas de ambas valvas.
La altura divide la concha en dos partes que pueden ser simétricas (concha equilateral), o asimétricas (concha inequilateral).
El cierre y apertura de las valvas se efectúa a través de la acción muscular. Sin embargo, las valvas están articuladas por unos dientecillos situados sobre plataformas conchíferas denominadas plataforma cardinal, localizada dentro de la concha en la región dorsal debajo de la zona que ocupa el músculo ligamentario. Estos dientes están separados por fosetas dentarias en las que encajan los dientes de la valva opuesta.
El cierre hermético de la concha queda asegurado en muchas conchas por la existencia de un conjunto de crenulaciones a lo largo del borde ventral de una de las valvas, que encajarían en los espacios intrecrenulares de la valva opuesta.
-
Tipos de dentición
El número y disposición de estos dientecillos sobre la plataforma cardinal tiene importancia desde el punto de vista sistemático. Se pueden distinguir vario tipos:
-
Taxodonta
Consiste en un conjunto bastante numeroso de dientecillos situados a lo largo de la plataforma cardinal y dispuestos simétrica o asimétricamente a ambos lados de los ganchos (parte más elevada de cada valva en la región dorsal).
-
Subtipo fenodonto
Las prolongaciones de estos dientecillos confluyen en un punto imaginario situado en el interior de la concha.
-
Subtipo pseudofenodonto
Las prolongaciones de los dientecillos confluyen en un punto imaginario fuera de la concha y más o menos próximo a su margen ventral.
-
Actinodonta
Conjunto de dientes de tamaño mayor que los anteriores, en número variable y dispuestos siempre sobre la plataforma cardinal e irradiando de los ganchos.
-
Heterodonta
Poseen dos tipos de dientes:
-
Cardinales
Se sitúan inmediatamente por debajo de los ganchos, en número de dos o tres, irradiando de ellos.
-
Laterales
Otros dos aparecen en las partes laterales de la plataforma cardinal.
-
Isodonta
Poseen dos o tres pares de dientes largos de tipo lamelar situados simétricamente a ambos lados de los ganchos y de una estructura que aloja el músculo ligamentario de tipo fibroso (foseta ligamentaria). También irradian de los ganchos.
-
Disodonta
Constituído por un conjunto de dientecillos en número variable, situados simétricamente a ambos lados de los ganchos y poco efectivos en cuanto a la articulación presente comúnmente en conchas de tipo mytiliforme (mejillón).
-
Musculatura
La musculatura de los lamelibranquios está constituída por dos tipos de músculos.
-
Músculos ligamentarios
Responsables de la apertura de la concha, en lamelibranquios actuales existen dos componentes musculares: lamelar y fibroso. Por su posición en la concha pueden ser:
-
Anfidéticos
La componente lamelar se sitúa simétricamente a ambos lados de la componente fibrosa.
-
Opistodéticos
La componente lamelar posee una magnitud de crecimiento mayor en la dirección posterior de la concha.
La disposición de ambas componentes (lamelar y fibrosa) es importante desde el punto de vista sistemático y provoca la distinción de varios tipos de ligamentos o músculos ligamentarios:
-
Duplivincular
Ligamento externo consistente en una serie alternante de capas lamelares y fibrosas que confluyen por debajo de los ganchos formando un ángulo más o menos agudo y que se insertan en la región dorsal de la concha.
-
Alivincular
La componente lamelar del ligamento se dispone simétrica o asimétricamente en la región dorsal a ambos lados de la componente fibrosa.
-
Alivincular anfidética
La componente lamelar se dispone de forma simétrica respecto del gancho.
-
Alivincular opistodética
La componente lamelar se dispone asimétricamente respecto al gancho con su crecimiento mayor en dirección posterior.
-
Multivincular
Formada por una serie de ligamentos alivinculares dispuestos en serie con un desarrollo bastante pronunciado hacia la región posterior de la concha.
-
Parivincular
El ligamento lamelar se sitúa siempre en la parte posterior de la concha alojado en un tubo cilíndrico por encima del ligamento fibroso.
-
Músculos aductores
Responsables del cierre hermético de la concha, una parte de éstos también aparecen relacionados con diferentes órganos del organismo, pero cuyas huellas sobre la concha en ocasiones son difíciles de interpretar, con la excepción de los músculos paleales, responsables de la inserción del manto a lo largo de todo el borde interior ventral de las valvas.
Dejan un par de huellas en el interior de la concha de forma redondeada y situadas en la región anterior y/o posterior unidas por la línea paleal correspondiente a los músculos paleales. Según el número de huellas existen:
-
Conchas monomiarias
Tienen una sola huella.
-
Conchas dimiarias
Tienen dos huellas.
-
Conchas dimiarias anisomiarias
Una huella es de tamaño muy superior a la de la otra.
-
Conchas dimiarias isomiarias
Las dos huellas son de igual tamaño.
-
Seno paleal
El manto puede verse dividido en determinados puntos por la aparición de sifones u órganos dependientes del manto y cuya función es la de inhalar o exhalar corrientes de agua. Las conchas provistas de sifones suelen dejar una marca muy clara, en la huella del músculo paleal, mostrando una discontinuidad por debajo de las huellas de los músculos, llamada seno paleal. Según esto, distinguimos dos tipos:
-
Intergripaleales
Este tipo de conchas poseen una línea paleal continua.
-
Sinupaleadas
Estas conchas tienen las conchas afectadas por un seno paleal.
-
Orientación de los ganchos
El ápice de los ganchos (parte más saliente de la región dorsal) de ambas valvas puede apuntar en distintas direcciones:
-
Prosogiros
Ambos ganchos apuntan hacia la región anterior.
-
Opistogiros
Ambos ganchos apuntan hacia la región posterior.
-
Ortogiros
Los ganchos miran el uno hacia el otro sin inclinación hacia ninguna región.
-
Enrollados
Los ganchos pueden estar enrollados, con el eje de enrollamiento dentro del eje de simetría comisural.
-
Espirogiros
El eje de enrollamiento está fuera del plano de simetría de la concha.
-
Ornamentación
-
Superficies diferenciales
El área exterior de la concha, aparte de su ornamentación particular, puede estar afectada por superficies diferenciales, cada una de las cuales posee una ornamentación diferente. Existen dos tipos de superficies diferenciales relacionadas con la región dorsal de la concha, denominadas lúnula y escudete, que tienen sección circular o elipsoidal, existiendo la lúnula siempre en la parte anterior de los ganchos, y el escudete en la posterior.
-
Carenas
Las carenas son un par de elevaciones considerables que pueden tener las valvas de un bivalvo. Están dispuestas hacia la región posterior de la concha, delimitando dos superficies externas de la concha claramente diferenciadas del resto en cuanto a su ornamentación. La parte anterior se llama área, y la posterior, corselete.
-
Otras ornamentaciones
Elementos radiales (estrías o costillas) o lamelas de crecimiento como elementos radiales; conjuntamente pueden existir prolongaciones de las valvas a modo de espinas relacionadas a adaptaciones a medios concretos. Algunas conchas viven fijas al sustrato a través de una de sus valvas, produciendo una cicatriz de cementación, fácilmente observable.
Criterios de orientación de la concha
En conchas de lamelibranquios actuales, la dirección del eje oroanal nos sitúa en las direcciones anterior y posterior de la concha. Dado que en lamelibranquios fósiles no nos es posible utilizar criterios en torno a las partes blandas, hemos de seleccionar otros criterios relacionados con estructuras fosilizadas que se sitúan de manera constante en las partes anterior y/o posterior de la concha.
-
En conchas dimiarias heteromiarias la huella del aductor de mayor tamaño está siempre dispuesta en la región posterior.
-
En conchas sinupaleadas, el seno paleal se sitúa siempre por debajo del aductor posterior.
-
El ligamento, con la excepción del alvincular anfidético, muestra siempre un desarrollo mayor hacia la región posterior de la concha.
-
La lúnula es siempre anterior y el escudete es siempre posterior.
-
Las carenas tienen un mayor desarrollo hacia la región posterior de la concha (si existen).
-
Los pectínidos poseen siempre un surco bisal próximo a la región dorsal de la concha situado en posición anterior y relacionado constantemente con la valva derecha.
A veces no se observan todas las características en la misma concha, pero siempre existe alguna.
Clasificación
Las clasificaciones actuales se basan en criterios anatómicos de las partes blandas. En paleontología no se puede y por ello acudimos a otros criterios representados por estructuras que fosilizan, de las cuales se hace especial mención al tipo de dentición, al tipo de musculatura y a la ornamentación.
En función de estas tres variables la mayor parte de los autores aceptan una clasificación en subclases: Paleotaxodontos, pteriomorfos, paleoheterodontos, criptodontos, heterodontos, anomalodermátidos.
4. Importancia
Los moluscos lamelibranquios aparecen en el Cámbrico inferior y empiezan a desarrollarse con importancia a partir del Ordovícico y Silúrico, donde ya aparecen todas las subclases. En el Devónico aparecen los primeros de agua dulce y en el Mesozoico y el Terciario hasta la actualidad aumenta su importancia, coincidiendo con la recesión de lo braquiópodos que ocupando los nichos que dejaron libres.
Tienen importancia paleoecológica puesto que son bastante sensibles a la salinidad y a la profundidad. En función de estas variables se sitúan las formas en distintos tipos de medios. También poseen interés estratigráfico, ya que evolucionan lentamente, salvo la excepción de los Rudistas.
-
Rudistas
Son un grupo de lamelibranquios heterodontos que aparecieron de manera súbita en el Jurásico superior y que sufrieron una extinción a finales del Cretácico, cuando las condiciones parecían las más ideales para seguir desarrollándose.
Son formas solitarias o bien agrupadas en pequeñas comunidades, llegando a formar bancos con ostreidos o con hexacoralarios.
Su esqueleto es de CaCO3, constituído por dos valvas inequivalvas como consecuencia de vivir fijo al sustrato por una de las valvas. La valva fija al sustrato toma un aspecto de cuerno más o menos retorcido o de cono más o menos regular. La valva libre puede tener una morfología similar (cuerno) o hacerse opercular, de morfología ligeramente cónica.
La dentición de los Rudistas es heterodonta paquiodonta, consistente en dos largos dientes en la valva libre entre los que existe una foseta y un solo diente en la valva libre con dos fosetas donde encajarían los dientes de la valva libre.
El ligamento es externo en formas primitivas e interno en las más modernas y en éstas está soportada por una arista ligamentaria que recorre longitudinalmente un surco ligamentario bastante profundo que recorre la valva fija.
La musculatura aductora es siempre dimiaria y se sustenta sobre dos apófisis calcáreas potentes llamadas Mióforos.
Poseen también estructuras sifonales soportadas por pilares sifonales relacionadas con la valva fija y por un par de aberturas u ósculos por donde saldrían estos sifones en la valva libre.
Los grupos más representativos de Rudistas aparecen en el Cámbrico superior y corresponden a hipurítidos y radiolitos. Con ellos se han establecido biozonas que son importantes y coinciden en general con algunas otras biozonas más importantes aún de ammonoideos.
Gasterópodos
Caracteres generales
Importantes paleontológicamente, son moluscos de concha dura que aparecen a partir del Cámbrico inferior y en el Ordovícico ya se presentan la mayoría de las subclases. Son capaces de habitar, marino, de agua dulce y terrestre.
Pierde ya en procesos tempranos de su desarrollo su simetría radial, como consecuencia de un proceso de torsión de las partes blandas, acompañado de un proceso de torsión de la concha.
En los ejemplares vivos, la cabeza está perfectamente desarrollada, poseyendo ojos, tentáculos, mandíbulas córneas y una placa rígida córnea o calcárea llamada rádula que ayuda en la masticación y en la predación.
El manto de los gasterópodos posee, al igual que el de los lamelibranquios, determinados repliegues que permiten la salida de los sifones con funciones exalantes e inhalantes de corrientes de agua hacia la cavidad paleal.
Comprenden animales acuáticos de agua dulce y salada, pero es el único filo de los moluscos que se ha logrado adaptar a la vida terrestre.
Mayoritariamente son formas bentónicas que se arrastran por medio del pie, aunque parcialmente o en algunos momentos de su vida pueden aparecer formas pelágicas.
-
Tipos de conchas
La concha es de CaCO3, externa y univalva. Generalmente arrollada en espiral, puede aparecer interna o muy reducida, pudiendo llegar a desaparecer totalmente.
-
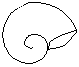
Formas de enrollamiento
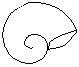
La concha puede ser visualizada originariamente como un tubo cónico, abierto por la región anterior o apertural y cerrado por la posterior o apical que se enrolla espiraladamente alrededor de un eje que pasaría por la región apical de dos formas:
-
Planispiraladas
El enrollamiento se produce siempre perpendicular al eje.
-
Conispiraladas
El enrollamiento se produce en espiral cónica, dado lugar a un cuerpo conispiralado, con la región apical más sobresaliente.
Cada circunvolución del tubo alrededor del eje constituye una vuelta, pero teniendo en cuenta que cada vuelta normalmente cubre de manera parcial a la precedente. A efectos prácticos, se considera como vuelta sólo la parte visible de cada una de las circunvoluciones.
-
Tipos de enrollamiento
Hay distintos tipos de concha en función del grado de cubrimiento de cada una de las vueltas sobre la precedente:
-
Concha convoluta
La última de las vueltas situada en la región anterior cubre e incluye totalmente al resto de las vueltas
-
Concha involuta
Cada una de las vueltas cubre parcialmente a la precedente.
-
Concha evoluta
Todas las vueltas se tocan tangencialmente.
El enrollamiento de la concha alrededor del eje puede producirse de manera muy apretada hasta el punto de que las partes internas de cada una de las vueltas situadas en la región axial pueden llegar a tocarse, generando una estructura calcárea que recorre axialmente toda la concha denominada columela. Cuando el enrollamiento es muy flojo las vueltas en la parte interna (axial) no llegan a tocarse, quedando un hueco de forma cónica que recorre también axialmente la concha denominada ombligo. Suele aflorar ceca de la apertura de la concha en su región anterior
-
Caracteres generales de la concha
-
Orientación
La orientación de la concha es tal que la dirección hacia la región apertural es anterior y la dirección hacia la región apical es posterior.
-
Aspecto general de la concha
Las primeras vueltas de cada una de las conchas suele tener una ornamentación y un enrollamiento generalmente diferente al del resto de la concha. Estas primeras vueltas corresponden a la protoconcha y el resto a la teleoconcha. Estas variaciones en el tipo de orientación y enrollamiento se deben a cambios bastante radicales en los hábitats de vida a lo largo del desarrollo de la concha.
Todo el conjunto de vueltas de un gasterópodo, con la excepción de la última, que es visible en toda su extensión, se denomina espira. La última vuelta de la concha es visible en toda su superficie y aflora anteriormente en una amplia abertura llamada peristoma, que puede ser:
-
Holostomado
Es continuo, sin ningún tipo de inflexión.
-
Sifonostomado
Está afectado por una escotadura sifonal en la región anterior extrema del peristoma.
En el peristoma que rodea la apertura se distinguen dos partes:
-
Labio externo
Equivale a la parte externa y abarca desde la línea de sutura hasta la región axial. Puede estar más o menos engrosado y poseer digitaciones variadas e incluso una escotadura en la región posterior extrema relacionada con la expulsión del deshecho, llamada escotadura anal.
-
Labio interno
Equivale a la parte interna y está constituída por dos porciones:
-
Porción columelar
Porción coincidente con la dirección del eje.
-
Porción parietal
Porción ligeramente oblicua al eje de la concha, que en muchas formas de gasterópodos posee depósitos secundarios de CaCO3 o de conquiolina (sustancia de naturaleza semicalcárea de aspecto nacarado) que se denomina callo o inductura.
-
Ornamentación
Su ornamentación es bastante variada, basándose en costillas, estrías, tubérculos, espinas, lamelas de crecimiento espesadas denominadas varices, cuya disposición responde a dos tipos:
-
Ornamentación colabral
Paralela al labio externo.
-
Ornamentación espiral
Paralela a la línea de sutura.
Algunas formas poseen opérculo, generalmente córneo, segregado por la parte posterior del pie y cuya función es tapar la apertura cuando la cabeza y el pie se retraen hacia el interior de la vuelta. Algunos son de CaCO3, por lo que se encuentran fósiles.
Clasificación
Los paleontólogos no han elaborado las clasificaciones de gasterópodos a un nivel mayor que el de familia porque la mayor parte de los fósiles evolucionan muy lentamente y se encuentran representados a lo largo de todo el registro geológico.
Es sencillo colocar las conchas de gasterópodos según las clasificaciones neontológicas, que se basan en caracteres anatómicos de las partes blandas. Para ello se utilizan los criterios de presencia o ausencia de determinados tipos de opérculos y la situación de la cavidad paleal y de las branquias. Según esto, hablamos de 4 subclases:
-
Protogasterópodos
Sólo fósiles, su concha no había experimentado torsión.
-
Prosobranquios
Cavidad paleal y branquias en posición anterior.
-
Opistobranquios
Cavidad paleal y branquias en posición posterior.
-
Pulmonados
Depósitos de branquias y la cavidad paleal convertidos en pulmón.
Interés
Aparecen en el Cámbrico inferior y su número aumenta a lo largo del registro geológico estando actualmente en expansión. De poca importancia estratigráfica, son bastante influenciables por las condiciones del medio, como salinidad y temperatura, por lo que tienen importancia paleontológica.
Moluscos camerados
Bajo este epígrafe se incluyen un grupo de moluscos de concha calcárea que internamente se encuentra tabicado por un número de septos más o menos grande que a su vez provocan una división de la concha en cámaras sucesivas. Dentro de ellos incluimos a los cefalópodos y a un pequeño grupo, el de los tentaculítidos, que presentan unas características que parecen demostrar que pueden incluirse dentro del grupo de los camerados.
Cefalópodos
1. Caracteres generales
Constituyen la clase más importante de los moluscos desde el punto de vista estratigráfico y desde el punto de vista evolutivo de los moluscos.
Representados por formas marinas y nadadoras, poseen simetría bilateral con respecto a un plano sagital que divide la concha en dos partes iguales, con la excepción de un pequeño grupo que representa el final de la línea filética concreta de ammonoideos: la simetría bilateral se pierde, haciéndose los ejemplares totalmente irregulares y agrupándose bajo la denominación de heteromorfos.
-
Concha
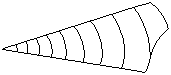
La concha de los cefalópodos es de CaCO3, univalva, externa, interna o pudiendo faltar totalmente y tabicada interiormente por septos transversales a la concha. En el único genero actual, nautilus, y en muchas formas conocidas fósiles, la concha externa y puede estar representada por un cono recto o puede encontrarse enrollada en un plano, planispiralada más o menos apretada. En el caso más sencillo, la concha aparece como un cono recto en el que aparecen dos partes diferenciadas:
-
Cámara de habitación
Se sitúa siempre en la parte anterior de la concha, ocupando de 1/3 a 1/4 del volumen total de la concha. En ella se alojan las partes blandas del animal que están constituidas por cabeza bien diferenciada, ojos muy similares a los de los vertebrados, boca rodeada siempre de tentáculos y provista normalmente de mandíbulas córneas y de rádula y un pie muy transformado en forma de embudo, con su parte más amplia abierta hacia la cavidad paleal, donde se encuentran las branquias.
-
Fragmocono
Se sitúa a continuación de la cámara de habitación e internamente está tabicado por un conjunto más o menos numeroso de septos transversales a la concha que lo dividen en cámaras. Cada uno de estos septos posee un agujero llamado foramen septal el cual puede estar continuado por un tubo de naturaleza conchífera llamado gollete sifonal.
Normalmente las partes blandas alojadas en la cámara de habitación tienen contacto directo con la cámara inicial a través de una prolongación carnosa relativamente fina llamada sifón, atravesando los septos por forámenes septales y golletes sifonales. Así, en los cefalópodos actuales el conjunto de estructuras relacionadas con el sifón que son los forámenes septales, golletes sifonales, el sifón y algún tipo de estructura mineralizada que los recubren y que unen los golletes sifonales con el septo siguiente llamados anillos conectantes constituyen el sifúnculo. El sifúnculo en formas fósiles queda representado exclusivamente por los golletes sifonales.
Esta estructura tiene bastante importancia en la orientación de conchas fósiles de cefalópodos puesto que se sitúa en una posición desplazada del centro, considerada siempre ventral.
La cámara inicial es la primera parte segregada por el animal y generalmente no se encuentra fósil, ni en formas rectas ni enrolladas, dada su fragilidad.
-
Morfología de la concha
Según el grado de enrollamiento tenemos:
-
Ortocónicas
En forma de cono recto.
-
Cirtocónicas
Ligeramente curvadas.
-
Ofiocónicas
Enrolladas más o menos apretadas. Dependiendo del grado de cubrimiento de cada una de las vueltas sobre la anterior, tenemos:
-
Evolutas
Las vueltas se tocan tangencialmente.
-
Convolutas
Cada vuelta cubre parcialmente a la que le precede.
-
Involutas
La última vuelta cubre totalmente a todas las anteriores.
-
Torticónicas
Son conchas ofiocónicas no planispiraladas.
Además, existen distintos tipos de enrollamiento:
-
Longiconos
Más largos que anchos.
-
Breviconos
Más anchos que largos.
-
Giroconos
Las vueltas no llegan a tocarse.
-
Orientación de la concha
La orientación de la concha en los cefalópodos es tal que la dirección hacia la cámara de habitación es anterior y la dirección hacia la cámara inicial es posterior.
En muchas conchas ortocónicas o incluso ofiocónicas en el margen de la apertura suele situarse un entrante cóncavo llamado seno hiponómico. Este marca la posición en la que esa concha tuvo el hiponomo, el cual se considera que está siempre en posición ventral, lo que proporciona un criterio absoluto para distinguir la zona dorsal de la ventral de la concha.
Cuando no existe seno hiponómico debemos recurrir al sifúnculo, el cual está siempre en posición ventral. En cefalópodos primitivos, el sifúnculo está casi siempre en posición ventral, lo que sirve para la orientación, pero en los evolucionados el sifúnculo está siempre en posición lateral.
-
Ornamentación
Es bastante variable y más recargada cuanto más evolucionado sea. En general, consiste en una ornamentación espiral siguiendo la línea de contacto entre las vueltas. Ornamentación transversal al margen de la apertura y manchas de color, correspondientes generalmente a la conquiolina.
-
Estructura de la concha
Las conchas que poseen su región ventral en la parte externa y más convexa de las vueltas se llaman conchas exogástricas, y las que la poseen hacia la parte interna, endogástricas.
La apertura de la concha en ocasiones puede estar fuertemente restringida por escotaduras de diferente morfología y estar asociadas a un opérculo córneo o calcáreo que puede llegar a fosilizar separadamente del propio ejemplar de cefalópodo.
Septos y golletes del interior de las conchas tienen una variabilidad en las distintas formas:
-
Septos
-
Procélicos
Los septos son cóncavos hacia la cámara de habitación.
-
Opistocélicos
Los septos son convexos hacia la cámara de habitación.
-
Golletes sifonales
Los golletes sifonales pueden estar o no desarrollados. Cuando están desarrollados estos pueden ser:
-
Ortocoanados
Rectos.
-
Cirtocoanados
Curvos.
Depende de hacia dónde estén orientados, podemos también clasificarlos:
-
Prosifonados
Orientados hacia la cámara de habitación.
-
Retrosifonados
Orientados hacia la cámara inicial.
-
Holocoanados
Los golletes sifonales llegan desde el foramen de un septo hasta el foramen del septo vecino.
Algunos moluscos cefalópodos poseen un esqueleto interno cuyas características coinciden parcialmente con las de los que tienen esqueleto externo (será abordado en los coleoideos).
2. Importancia
Aparecen en el Cámbrico inferior y se desarrollan con diferente importancia a lo largo del tiempo. La mayor parte de las formas con esqueleto externo se han extinguido, salvo el género nautilus, en el que hay diferentes especies. Las formas de esqueleto interno han ido progresando y en la actualidad están muy representadas.
3. Línea de sutura
Es el rasgo fundamental, básico en la sistemática y en la filogenia de los cefalópodos, y muy especialmente la de los ammonoideos.
Es una línea más o menos alabeada que resulta de la intersección de los septos con la concha. Normalmente, esta intersección está enraizada en las capas más internas de la concha. Para su estudio es necesario levantar la capa más externa de la concha (periostraco), e inclusive desgastar la segunda (ostraco). El estudio de la línea de sutura debe realizarse a lo largo de la ontogenia, a media que el cefalópodo se va desarrollando.
Son de considerar especialmente la línea de sutura que deja el primero de los septos nacidos, que delimita la cámara inicial y el resto del fragmocono, que se denomina prosutura. En segundo lugar, la evolución de la línea de sutura durante los tres o cuatro primeros septos nacidos a continuación del primero, que constituyen la sutura primaria. Por último, la evolución final de la línea de sutura correspondiente a los últimos septos más próximos a la cámara de habitación, que constituyen la sutura adulta.
En formas enrolladas de cefalópodos, la línea de sutura de cada septo se desarrolla tanto en la parte externa de las vueltas como en la interna, por lo que para su estudio detallado es necesario ir despejando sucesivamente todas las vueltas para que las líneas de sutura sean observables en su totalidad.
La parte de la zona de sutura correspondiente a la zona externa de la vuelta se llama sutura externa, y la correspondiente a la parte interna, sutura interna.
A efectos descriptivos, la línea de sutura está constituída por partes cóncavas hacia la cámara de habitación (lóbulos), y partes convexas hacia la cámara de habitación (surcos o sillas). Cada uno de estos elementos suturales se simbolizan con una letra mayúscula a veces acompañada de un subíndice numérico. El símbolo indica el tipo de elemento sutural y el índice el orden de aparición. El conjunto de todos estos símbolos correspondientes a elementos suturales dispuestos desde la región ventral a la dorsal recibe el nombre de fórmula sutural. Normalmente estas fórmulas se efectúan sólo con los símbolos de los lóbulos, habida cuenta que entre cada dos lóbulos ha de existir necesariamente el correspondiente surco.
Los surcos no suelen simbolizarse, pero en el caso de que fuese necesario distinguir alguno de hace con las letras de los símbolos de los lóbulos adyacentes, poniendo en primer término el del lóbulo más próximo al área ventral y en segundo término el más alejado.
Las suturas pueden representarse gráficamente en un plano indicando sobre ellas la traza del plano de simetría mediante una flecha cuya dirección indica hacia donde está la cámara de habitación. La flecha de mayor tamaño indica la parte más convexa y externa de la vuelta, y la de menor tamaño, la parte interna de la vuelta.
En el caso de mayor complicación sutural, aparecen una serie bastante numerosa de lobulillos que describimos a continuación:
Los tres primeros lóbulos que aparecen corresponden a la sutura primaria, y se denominan externo, interno y lateral.
Aparecen también lóbulos secundarios que juntamente con los primarios determinan la sutura adulta: adventicios, umbilicales y accesorios.
-
Lóbulos primarios
-
Lóbulo externo (E)
Es el lóbulo primario situado en la parte externa de la concha y en su región más convexa. Dividido por la traza del plano de simetría, se simboliza por la letra E. En ocasiones, en su fondo puede ocurrir una pequeña inflexión en cuyo techo se produce un nuevo lóbulo de pequeño tamaño, llamado lóbulo medio.
-
Lóbulo interno (I)
Es el lóbulo primario situado en la parte cóncava (interna) de las vueltas. Dividido por la traza del plano de simetría, se representa por la letra I.
-
Lóbulo lateral (L)
Es el primero de los lóbulos de la concha que aparece en los flancos, entre el externo y el interno. A veces, ocupa todo el flanco. Representado por L.
-
Lóbulos secundarios
Una vez que han aparecido los lóbulos que determinan la sutura primaria, aparecen los lóbulos secundarios, que nacen de los surcos que separan los lóbulos primarios.
-
Lóbulos adventicios (A)
Son los lóbulos secundarios que nacen a partir del techo del surco que separa los lóbulos externo y lateral. Representados por la letra A, pueden ser varios y llevar subíndices según el orden de aparición: A1, A2, A3,...
-
Lóbulos umbilicales (U)
Son los lóbulos secundarios que aparecen a partir del surco que separa los lóbulos lateral e interno. Se representan por U y, al igual que los adventicios, pueden existir varios, llevando subíndices según el orden de aparición.
-
Lóbulos accesorios (K)
Son de muy pequeño tamaño y terminan recortando tanto surcos como lóbulos generados anteriormente. Se denominan por la letra K, afectan a todos los lóbulos anteriormente nacidos y son muy numerosos.
Algunos autores, sobre todo rusos, han tratado de proponer alternativas de símbolos diferentes a éstos, llamando por ejemplo ventral al externo, y al interno, dorsal, y también para los lóbulos secundarios, pero han caído en desuso, siguiendo la mayor parte de los especialistas la nomenclatura alemana.
Normalmente la línea de separación de la sutura externa e interna se suele marcar por una línea de trazos. Las líneas de sutura se hacen con la mitad de la estructura ya que la otra es idéntica (son simétricas).
4. Clasificación
El género actual nautilus y con toda probabilidad, por aplicación del Principio del Actualismo las restantes conchas fósiles externas de cefalópodos, poseyeron dos pares de branquias, mientras que los cefalópodos actuales, con esqueleto interno o sin él, así como un numeroso grupo de fósiles de cefalópodos correspondientes a los coleoideos, poseen un solo par de branquias. Por ello se clasifican en Neontología en tetrabranquios y dibranquios.
Esta clasificación no puede aplicarse en paleontología, por lo que hemos de acudir a otros criterios esqueléticos basados solamente en la línea de sutura, tanto la prosutura como la sutura primaria, la posición del sifúnculo y la disposición de los golletes sifonales. En función de estas características, Teichert en 1967 propuso una clasificación según las siguientes subclases:
-
Nauliloideos
-
Bactritoideos Sólo fósiles, con concha externa
-
Ammonoideos
-
Coleoideos Amplia representación actual. Esqueleto interno
-
Nautiloideos
Grupo sencillo de cefalópodos que básicamente está constituído por cuatro órdenes diferentes: ortocerátidos, actinocerátidos, endocerátidos y nautiloideos.
Poseen concha de CaCO3, ortocónica, cirtocónica o planispiralada. Ornamentación sencilla, basada en líneas de crecimiento exclusivamente. En la prosutura la concavidad del primero de los tabiques es siempre procélica.
La sutura primaria no tiene lóbulos en formas poco evolucionadas, aunque en formas muy evolucionadas puede llegar a desarrollar los tres lóbulos primarios.
El sifúnculo siempre en posición subventral o más o menos variable y los golletes sifonales son retrosifonados.
Aparecen en el Cámbrico superior y llegan hasta la actualidad.
-
Bactritoideos
Grupo poco evolucionado que es origen de los ammonoideos. Sus características ya parecen preludiar las más sencillas de formas evolucionadas.
Son conchas ortocónicas o cirtocónicas de pequeño tamaño (2,3 cm) con prosutura recta o ligeramente cóncava. Su sutura primaria está constituida por un lóbulo externo y dos lóbulos laterales. Sifúnculo constantemente en posición ventral, pero no siendo siempre su disposición retrosifonada.
Aparecen en el Ordovícico y se extinguen en el Pérmico. Son sólo doce géneros.
-
Ammonoideos
-
Caracteres generales
Es el grupo más importante de cefalópodos desde el punto de vista estratigráfico. Aparecen en el Devónico superior y se extinguen a finales del Cretácico (piso maastrichtense).
Concha de CaCO3 fuertemente tabicada (camerada) con un tamaño de 2 cm hasta formas gigantescas de hasta casi 2 m de diámetro. Siempre es externa, enrollada planispiraladamente.
La terminología de enrollamiento es igual en general salvo unas excepciones:
-
Oxiconos
Formas lenticulares de bordes cortantes y fuertemente involutas.
-
Esfericonos
Concha de bordes externos redondeados y también involutas.
-
Serpenticonos
Formas evolutas
-
Crioconos
Formas cuyas vueltas están despegadas.
La prosutura de los ammonoideos es siempre opistocélica, y la sutura primaria puede ser bastante sencilla en formas primitivas, pero se complica progresivamente a lo largo de la evolución, tanto en la ontogenia como en la filogenia del grupo
Según su morfología se diferencian dentro del grupo tres tipos de suturas:
-
Goniatítica
Todos o la mayoría de los lóbulos no están subdivididos, o lo están pobremente.
-
Ceratítica
Posee siempre lóbulos dentados o serrados y los surcos son lisos o redondeados.
-
Ammonítica
Tanto lóbulos como surcos están fuertemente afectados por lobulillos de pequeño tamaño de tipo accesorio.
El sifúnculo está siempre en posición ventral. Las formas primitivas son retrosifonadas al igual que las formas primitivas de bactritoideos y nautiloideos. Las formas más evolucionadas son siempre prosifonadas.
La ornamentación es tanto más recargada cuanto más evolucionado sea. Constituida por multitud de elementos ornamentales tales como estrías, costillas, quillas, prolongaciones diversas de tipo tubérculo o espina, etc.
Hay placas operculares en algunas formas que tapan la apertura y que son calcáreas, por lo que pueden fosilizar.
Su prosutura es opistocélica.
-
Clasificación
Basándose en el análisis y observación de la línea de sutura, tanto la prosutura, como la adulta, la observación de la posición del sifúnculo y de los golletes sifonales, se distinguen ocho órdenes distintos agrupados en tres conjuntos cuyos nombres hacen referencia al período en el que han vivido.
| | Paleoammonoideos | Mesoammonoideos | Neoammonoideos |
| Etapas | Durante el Paleozoico, del Devónico inferior al Pérmico | Un sólo orden en el Pérmico; durante el Triásico mayor desarrollo | Proceden de los Mesoammonoideos. Durante Triásico y Cretácico |
| Órdenes |
|
|
|
| Sutura primaria | Goniatítica | Ceratítica | Ammonítica |
| Fórmulas suturales | Trilobada (Externo, interno y lateral) | Sutura primaria: 4 lóbulos (externo, interno, lateral y umbilical) | * |
| Suturas adultas: Muy complejas. Abundantes lóbulos accesorios. |
*Neoammonoideos: presentan lóbulos bífido y cruciforme. Según el orden se caracterizan por:
Filocerátidos: Sutura adulta quincuelenada. Presentan la sutura más ammonítica de todo el grupo.
Ammonítidos: Hay más lóbulos en la sutura primaria que en la adulta. Los lóbulos pasan a ser surcos (regresión sutural). Pueden pasar de ser de sutura ammonítica a goniatítica o ceratítica.
Litocerátidos: En el Cretácico superior tienen su mayor apogeo, perdiendo la simetría, generándose formas llamadas heteromorfas (en este grupo es donde hay más). Es debido a que hay gran desarrollo, empiezan a desarrollarse tanto que las conchas llegan a ser totalmente irregulares, hasta turriculadas (parecidas a las de los gasterópodos).
Coleoideos
Caracteres generales
Comprenden los cefalópodos actuales con concha interna o sin concha, y un conjunto de formas fósiles cuyas peculiaridades esqueléticas sugieren que su esqueleto podía ser interno.
Los esqueletos son de CaCO3 en general y están compuestos por un fragmocono más o menos alargado fuertemente tabicado por septos transversales, que en posición anterior muestran una cámara de habitación prolongada en una estructura dispuesta a modo de visera denominada prostraco. Posteriormente, el fragmocono está incluido en una pieza maciza en forma de bala denominada rostro que normalmente es lo único que aparece fósil.
Aparecen durante el Carbonífero, y muchas de sus formas se extinguen al final del Cretácico, pero otras llegan a la actualidad donde están representadas ampliamente en los fondos marinos.
2. Clasificación
-
Aulacocerátidos
-
Fragmoteútidos Sólo fósiles
-
Belemnítidos
-
Teútidos
-
Sépidos Representados en la actualidad
-
Octópidos
-
Belemnítidos
Grupo de fósiles más importante. Su fragmocono es en general corto, con un sifúnculo en posición siempre ventral, prostraco relativamente largo y situado en la región dorsal. Poseen una pieza adicional llamada epirostro situada en posición posterior que es donde encaja el rostro de forma más o menos similar a como lo hace el fragmocono con respecto al rostro.
Normalmente suelen encontrarse fósiles tanto el rostro como el epirostro porque son estructuras macizas que se desprenden del fragmocono y que pueden encontrarse diseminadas entre los sedimentos.
Aparecen en el Jurásico, y se extinguen a finales del Cretácico excepto un género que se adentra un poco en el Terciario
-
Representantes actuales
Los Sépidos se caracterizan por tener un prostraco muy fuerte, y los Téutidos son pulpos gigantes, de hasta 18 metros.
3. Importancia general de los cefalópodos
Dentro de los cefalópodos están los fósiles más importantes desde el punto de vista cronoestratigráfico del registro debido a dos razones:
-
Tienen la capacidad de natación y por ello una gran dispersión geográfica (cosmopolitas), apareciendo en casi todas las cuencas paleozoicas y mesozoicas del mundo.
-
Las especies poseen una rápida evolución y por ello se pueden establecer biozonas estratigráficas; son por ello buenos fósiles guía.
Los grupos más primitivos se desarrollan a lo largo del paleozoico: los Nautiloideos esencialmente en el Paleozoico inferior y medio y los Bactritoideos en el paleozoico superior.
A partir del Paleozoico los Nautiloideos bajan en número y se hacen menos importantes frente al creciente aumento de los Ammonoideos que al extinguirse estos, vuelven a adquirir cierta importancia numérica para volver a remitir posteriormente. Actualmente están en vías de extinción, representados sólo por un género (Nautilus), con sólo cinco especies.
Los Bactritoideos tienen poca importancia numérica y no han sido utilizados con aplicaciones cronoestratigráficas; sin embargo, tienen importancia desde el punto de vista filogenético porque actualmente se cree que a partir de ellos se originaron los cefalópodos evolucionados, tanto Ammonoideos como Coleoideos.
La posesión de la línea de sutura de los Bactritoideos de un lóbulo externo y dos laterales preludia la sutura de los Ammonoideos más primitivos, los Anarcétidos, Aunque la disposición constantemente ventral del sifúnculo hace pensar que ellos podrían haber derivado de los Coleoideos por la pérdida del esqueleto externo, pasando a ser interno o a faltar totalmente.
Dentro de los Ammonoideos están los principales fósiles guía de las biozonas que caracterizan el periodo de tiempo en el que han vivido desde el Devónico inferior al Cretácico superior. Representan los fósiles más importantes cronoestratigráficos del registro paleontológico, hasta el punto de que sus formas han sido consideradas junto con los graptolitos como ortocronológicas, a las que tienen que referirse cualesquiera otros tipos de correlación paleontológica.
Los Mesoammonoideos están representados sólo por el orden de los Ceratítidos. Comienzan su historia en el Pérmico pero su importancia la tienen en el Terciario a final del cual se extinguen. A lo largo del Triásico existen más de 400 géneros de ceratítidos que han proporcionado aproximadamente 30 biozonas. Teniendo en cuenta que el Triásico dura aproximadamente 25 m.a. cada biozona representa un lapso temporal de unos 800 a 850.000 años. Este lapso está próximo del poder máximo de resolución de la correlación estratigráfica.
Los Neoammonoideos se desarrollan durante el Jurásico y el Cretácico esencialmente, aunque algunos aparecen ya en el Triásico. El grupo más importante, el de los Ammonítidos, del que se han descrito más de 800 géneros (Jurásico - Cretácico) y que se conocen alrededor de 87 biozonas; el lapso temporal medio de cada biozona representa aproximadamente 1 m.a. para el Jurásico y 2 m.a. para el Cretácico.
Los Coleoideos aparecen en el Carbonífero; algunos grupos como los Belemnítidos tienen interés numérico en el Cretácico pero se extinguen rápidamente a principios del Terciario. Otros grupos han llegado hasta la actualidad, donde son mejor conocidos.
La desaparición de los Ammonoideos a finales del Cretácico superior es un problema de difícil interpretación dado que a finales de esta época existían unos 35 géneros de ammonoideos sin ningún tipo de debilitamiento, aunque esta debió ser repentina en ese mismo momento.
Se puede decir que la vida de los cefalópodos transcurrió a través de fuertes crisis en determinados momentos, en las que la supervivencia general del grupo dependió de la pervivencia de un sólo género o de una única línea filética. Fueron varias las crisis pero las más importantes que afectaron también a numerosos grupos de invertebrados y de vertebrados tuvieron lugar a finales del Pérmico y a finales del Cretácico (entre Cretácico y Terciario). La crisis del Pérmico fue la más importante porque desaparecieron casi el 95% de las especies de invertebrados marinos y también especies de vida terrestre.
Tentaculítidos
Caracteres generales
Constituyen un grupo de formas extinguidas, exclusivamente marinas. Su esqueleto es calcáreo en forma de cono recto de pequeñas dimensiones (máximo 2 - 3 cm) abiertos al exterior por la región anterior y desprovistos de opérculos.
Su simetría es aparentemente radial, pero desde hace pocos años algunos especialistas han descubierto huellas musculares en el interior de la concha dispuestas simétricamente respecto a un plano de simetría sagital o longitudinal.
-
Ornamentación
Es bastante sencilla, basada en estrías o costillas longitudinales y anillos más o menos espesos dispuestos transversales. Pueden aparecer proyecciones variadas (tubérculos) y excepcionalmente pueden ser lisos.
-
Partes de la concha
La concha puede dividirse en tres partes:
-
Región apical
Cámara inicial o embrionaria siempre, los anillos se disponen a espacios más o menos regulares, poseyendo el mismo espesor.
-
Región media
Los anillos poseen espesor variable y están separados por distancias bastante irregulares.
-
Región apertural
Los anillos son más finos que en el resto de la concha y están muy próximos unos a otros, separados por espacios muy regulares. En la parte terminal de esta región aflora la apertura.
Clasificación
En función de la morfología de la cámara inicial y del tipo de ornamentación de la concha se consideran tres órdenes:
-
Tentaculítidos
Cámara inicial cónica y alargada. El resto de la concha está ornamentada sólo por anillos. Una sección perpendicular a la cámara inicial muestra la presencia de 2 ó 3 tabiques espesos que delimitan cámaras por lo que algunos autores creen que pertenecen al grupo de los camerados. Las conchas son de 2 - 3 cm.
-
Darioconáridos
Formas microscópicas, conchas milimétricas, cámara apical subesférica o en forma de gota de agua y ornamentación constituida por elementos longitudinales y transversales que le confieren un aspecto reticulado son sus principales características.
-
Homocténidos
Poseen cámara inicial abombada con una prolongación posterior muy fina y el resto de la concha con ornamentación de anillos transversales.
Importancia
Los tentaculitoideos aparecen en el Ordovícico y se extinguen a finales del Devónico, casi en el límite entre los dos pisos del Devónico superior. Este grupo tiene importancia paleontológica:
-
En los tentaculítidos, paleoecológica porque son formas bentónicas que aparecen en plataformas someras acompañados de otras formas bentónicas (corales, braquiópodos, etc.)
-
Los Darioconáridos tienen interés estratigráfico a lo largo del Devónico inferior y medio, puesto que son formas pelágicas y sus especies evolucionan con rapidez. Están relacionados con facies profundas y alejadas de la costa.
Apertura
![]()
Escotadura
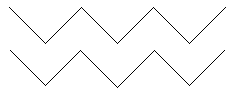
Dientes
Fosetas
Eje
Conchas planispiraladas
Conchas conispiraladas
![]()
Ortocónicas
![]()
Cirtocónicas
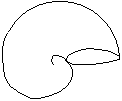
Involutas
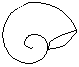
Convolutas
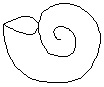
Evolutas
![]()
Torticónicas
Ofiocónicas
Longiconos![]()
Breviconos
![]()
Giroconos
Prostraco (cámara de habitación)
Fragmocono
Rostro (sólo fósil)
![]()
Descargar
| Enviado por: | Andrés Rodriguez |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.