Biología, Botánica, Genética y Zoología
Micorrizas
MICORRIZAS
Aspectos fisiológicos de la simbiosis
Universidad Nacional de Colombia
Resumen. El término de micorriza se aplica a la asociación simbiótica de plantas con algunos hongos del suelo. Las micorrizas en sus diferentes asociaciones cumplen un papel importante en el desarrollo y crecimiento de las plantas hospedantes; las asociaciones están directamente influenciadas dependiendo la especie de la planta y el hongo, el tipo de suelo y su interacción. Se dividen en Ectomicorrizas (EC), Ectendomicorrizas y Endomicorrizas, en estas últimas las más importantes son las micorrizas vesiculo-arbusculares (VAM), y en cuanto a su uso como biofertilizantes, en este trabajo se tratan aspectos referentes a la absorción, almacenamiento y transporte de P y N por plantas micorrizadas, y algunos ejemplos de micronutrientes tales como Zn, Cu, Mn y Fe, al tiempo que se resalta su impacto en el desarrollo de la planta.
Palabras claves: Micorriza, transporte, nutrición.
INTRODUCCIÓN
En condiciones naturales, los hongos del suelo colonizan las raíces de ~90% de las plantas terrestres formando micorrizas, asociación simbiótica que juega un papel importante en el ecosistema capturando los nutrientes del suelo. Más de 6000 especies de hongos son capaces de establecer asociación micorrizógena con casi 240000 especies de plantas, pero relativamente pocas interacciones planta-hongo son eficientes. Las plantas micorrizadas juegan a un precio - entre 10 y 20% de la fotosíntesis neta es requerida para la formación, mantenimiento y función de las estructuras de la micorriza-. Las asociaciones micorriza-arbúsculo (AM) son el tipo más común, que ocurre en ~80 especies de plantas (Jakobsen & Rosendahl, 1990; Bonfante & Perotto, 1995; Ayling, et al., 1997)
La infección por micorrizas puede proteger las raíces de la planta de patógenos del suelo, y quizá también incrementar el crecimiento de las raíces y la toma de los nutrientes por las raíces hospedantes. El mayor efecto sobre la nutrición es el resultado del transporte de la hifa de iones minerales poco o nada móviles. Las hifas crecen más allá de la rizosfera del suelo incrementando la superficie absorbente de las raíces. La actividad del micelio de AM resulta en un incremento en la eficiencia en la absorción de nutrientes. Este proceso es particularmente importante para la difusión lenta de iones minerales tales como el fósforo (P) (Smith & Gianinazzi-Pearson, 1988; Subramanian & Charest, 1999).
Solamente los tejidos específicos de raíces tales como tejidos epidermales y corticales son colonizados, mientras que los meristemos o tejidos vasculares son resistentes a la infección por micorrizas (Bonfante & Perotto, 1995). La simbiosis depende de los factores ambientales y en particular de la capacidad del hongo para colonizar las raíces y su funcionalidad en la toma de nutrientes. A este respecto, es importante diferenciar entre las Ectomicorrizas (ECM), Ectendomicorrizas y Endomicorrizas (Marschner & Dell, 1994).
1. TIPOS DE ASOCIACIONES
1.1 eCTOMICORRIZA (ECM)
Son asociaciones mutualistas entre el hongo y plantas de Gimnospermas o Angiospermas. Se desarrolla sobre la raíz una vaina o manto micótico (espesor 20 - 100 µm); y la penetración de la corteza por el hongo es intercelular. Se suprime la formación de pelos radicales y, en su lugar, se estimula la formación de finas raíces con micorrizas que se ramifican, en ocasiones, repetidamente. Los hongos que las forman son, principalmente, Basidiomicetos (cerca de 4000 especies) y unos pocos Ascomicetos (Sylvia, 2000).
Algunas hifas crecen entre las células de la epidermis y corteza radical formando la denominada “red de Harting” (Figura 1), que proporciona una gran superficie para los intercambios de materiales entre el huésped y la planta. Aquel recibe de la planta los compuestos carbonados y sustancias específicas de crecimiento, como la tiamina.
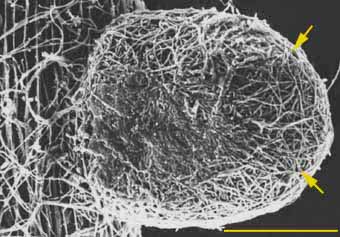
Figura 1. Colonización en Pino por Pisolithus tinctorius. Se observa el manto (flechas) formado por una vaina de hifas formada por una ectomicorriza. (Bar = 100 um) (Yves Piché & Larry Peterson)
La vaina micótica que se forma alrededor de las raíces proporciona mayor superficie de absorción de los compuestos minerales que la de la raíz sola, lo que favorece el crecimiento de la planta en suelos de baja fertilidad. Los fosfatos absorbidos son transportados más lentamente a los brotes de las plantas con micorrizas, por lo que se ha sugerido que debe producirse una acumulación en las vainas antes de ser transferidos al anfitrión (Wild, 1992).
Las Ectomicorrizas se encuentran en un rango amplio de arbustos y árboles forestales. Muchos de los hospedantes pertenecen a las familias Pinaceae, Fagaceae, Betulaceae y Myrcataceae. Algunos hongos tienen un angosto rango de hospedantes, tales como Boletus betulicola sobre Betula spp., al tiempo que otros tienen un amplio rango de hospedantes, tales como Pisolithus arhizus (también llamados P. tinctorius) el cual forma ectomiccorriza con más de 46 especies de árboles pertenecientes como mínimo a ocho géneros (Sylvia, 2000).
1.2 ECTENDOMICORRIZAS
En este tipo de micorriza concurren las características de las ectomicorrizas al tiempo que hay penetración al interior de las células corticales por hifas septadas. Estos hongos se encuentran asociados con coníferas, principalmente Ericales. Los hongos asociados son reconocidos como formadores de ectomicorrizas, lo cual se convierte en un argumento para que esta asociación se considere como una caso especial de este tipo de simbiosis.
Las ectendomicorrizas por su especificidad en Ericales podrían se consideradas poco importantes, pero en la medida que se estudian, se encuentra que participan en la conservación de los ecosistemas naturales (Sánchez, 1999).
1.3 ENDOMICORRIZAS
Entre las endomicorrizas se agrupan algunos tipos muy específicos como los correspondientes a las orquídeas y ericoides, y el más extendido dentro de las especies vegetales, la endomicorriza vesiculo-arbuscular (VAM). Estas últimas son simbiontes obligados que tienen una relación estructural intima con su planta hospedante, la cual involucra modificaciones de la estructura y composición de la pared celular de ambos patrones. Entre la raíz de la planta y el hongo puede tomarse un número de formas distintas de transferencia de nutrientes: hifa intercelular e intracelular y arbúsculo intracelular. Su crecimiento se produce característicamente en el interior de la raíz, no forman vaina y el micelio externo es, relativamente, escaso (Bonfante-Fasolo & Grippiolo, 1982; Wild, 1992; Avlung, et al., 1997).
Se han encontrado VAM en las raíces de numerosas plantas, incluso entre las que forman ectomicorrizas. La intensidad de la infección es variable presentándose casos, como en la familia Cruciferae, en que son muy débiles o no existen. Las VAM son el resultado de la colonización de raíces jóvenes por hongos de la familia Endogonaceae; de ella, los géneros Gigaspora, Glomus, Acaulospora y Sclerocystis son las más importantes (Wild, 1992).
La infección se inicia con las hifas, procedentes de una espora en germinación, formando un apresorio sobre la superficie de la raíz, y penetrando desde éste en las células epidérmicas. Las hifas, dispersas inter e intracelularmente a través de la corteza externa, proliferan en las capas corticales más internas formando arbúsculos (Figura 2). Un arbúsculo se desarrolla dentro de la célula infectada por ramificación dicotómica repetida de la hifa invasora. Los arbúsculos tienen vida corta y se desintegran en el suelo cuando muere la raíz.
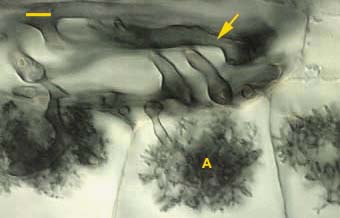
Figura 2. Raíces de Asarum canadense. VAM, en el género Glomus produce Arbusculos (A), hifas
Después de la ramificación arbuscular, las estructuras vesiculares se desarrollan como protuberancias inter e intracelulares a lo largo, o en el extremo, de las hifas. Sus esporas tienen altos contenidos de lípidos (cerca del 45% de su peso seco). Los más abundantes son triglicéridos, el contenido de fosfolípidos es bajo y ácidos grasos. Las esporas e hifas producen auxina, giberelina y citoquininas. Si estas son secretadas durante la formación de la micorriza, ellas puede contribuir a las cambios en la fisiología del hospedante (p. ej. La auxina estimula la actividad de las bandas de las membranas de ATPasa). VAM no puede penetrar la endodermis o cualquier pared que contenga suberina y lignina, indicando que esta no puede degradar estos componentes. El tubo germinal absorbe fosfato (P) por un proceso sensible a la temperatura, pero los mecanismos de toma y flujo no han sido ampliamente estudiados (Smith & Gianinazzi-Pearson, 1988; Wild, A. 1992; Bonfante & Perotto, 1995).
La intensidad de la infección por VAM puede estar regulada por varios factores: nutrición de la planta y fertilización, pesticidas, intensidad luminosa, humedad del suelo, pH y susceptibilidad de la planta. Como el hongo crece, la membrana celular del hospedante se invagina y es envuelta por el hongo, creando un nuevo compartimento donde el material de alto peso molecular es depositado. El espacio apoplástico previene el contacto directo entre la planta y el citoplasma del hongo y permite la transferencia eficiente de nutrientes entre los simbiontes. Los arbúsculos tienen relativamente vida corta, menos de 15 días (Wild, 1992; Sylvia, 2000).
1.3.1 Micorriza en orquídeas
Las orquídeas pasan por un estado de semillero prolongado durante el cual son incapaces de fotosintetizar, además que éstas tienen escasas reservas de almidones y lípidos. En condiciones naturales, estos requerimientos son suplidos por el hongo micorrizógeno. La planta es colonizada después de la germinación, y el hongo suple el carbono y las vitaminas para el desarrollo del embrión hasta cuando se forman las hojas y se inicia la fotosíntesis (Figura 3). Los hongos que participan en la simbiosis son basidiomicetos, algunos patógenos como Armillaria, Fomes, Marasmius y Rhizoctonia (R. Solani y R. Repens) (Gianinazzi-P & Gianinazzi, 1983; Sylvia, 2000).
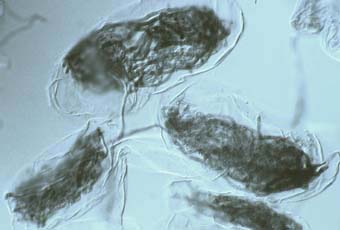
Figura 3. Micorrizas en orquídeas. Consiste de enrollamientos de la hifa ente las raíces o tallo de la planta
1.3.2 Micorriza Ericacea
El término ericaea es aplicada a asociaciones micorrízicas encontradas en plantas en el orden Ericales. Las hifas en las raíces pueden penetrar células corticales (hábitat de las endomicorrizas), sin embargo, no hay formación de arbúsculos (Sylvia, 2000).
2. ASPECTOS FISIOLÓGICOS DE LA SIMBIOSIS
La toma de nutrientes de la planta está determinada principalmente por la capacidad de absorción de las raíces y por la difusión de los nutrientes y subsecuente liberación de elementos en la solución del suelo. La tasa de absorción de iones con alta movilidad, tal como NO3-, de la solución del suelo es específica del cultivar y la especie. La capacidad de toma de iones con baja velocidad de difusión, p. ej. Zn y Mo y en menor grado K, S y NH4+, depende de la densidad de la raíz por volumen de suelo. En estos casos, la morfología de la raíz y el micelio externo del hongo VAM determina la tasa de toma de nutrientes para la planta (Sieverding, 1991).
2.1 DESARROLLO Y ESTRUCTURA PLANTA-HONGO EN RELACIÓN CON EL TRANSPORTE de nutrientes
A nivel celular, la interfase en todos los tipos de micorrizas están compuestos de las membranas de ambos patrones, separados por una región del apoplasto (Smith & Smith, 1990). El desarrollo intracelular afecta la estructura de la pared del hongo, el metabolismo y la fisiología, incluyendo la represión de la superficie y enzimas extracelulares (Smith & Gianinazzi-Pearson 1988). La reacción de penetración y proliferación del hongo parece similar en diferentes endomicorrizas en la presencia del haustorio intracelular; pero las células del hospedante muestran modificaciones más específicas. En todos los casos, la interfase formada entre la célula de la planta y el hongo involucra una nueva formación de la membrana de la planta que se extiende desde la membrana plasmática de la periferia alrededor del haustorio del hongo (Smith et al., 1994).
En orquídeas, el material de la pared celular persiste alrededor de la extensión intracelular. La presencia de pectinas y celulosa constituye la interfase arbuscular, simultáneamente con la actividad neutral fosfatasa indica que el protoplasto de la planta conserva la actividad de sintetizar la pared celular. La transferencia de carbohidratos de micorrizas en orquídeas es del hongo a la planta y sintetiza los componentes de la pared, no contribuye a la nutrición del hongo. La modificación en términos de transporte de nutrientes es la actividad de ATPasa en la membrana de la planta formada alrededor de los arbúsculos. En VMA parte de esta actividad es atribuible a un ion H ATPasa presente en la membrana periarbuscular, pero citoquímicamente indetectable a lo largo de otras membranas de la planta adyacentes a la hifa. La distribución de la actividad alrededor del arbúsculo sugiere que las células de las raíces infectadas tienen la capacidad de incrementar la absorción de nutrientes (Marx, et al., 1982; Smith, 1994).
En ectomicorrizas, los cambios estructurales en el tejido de la planta es pequeño, pero en algunos casos el crecimiento de la pared hacia dentro, parece ser inducida por la infección del hongo. La actividad ATPasa está asociada con la membrana plasmática de ambos simbiontes cuando estos son activos y estrechamente asociados en la región “Harting”, pero esta desaparece en células de plantas senescentes (Smith, 1994). La información de ultra estructura para todos los tipos de micorrizas indica que la acumulación de material en la interfase es reducida, en términos de la permeabilidad del apoplasto. Sin embargo, el material extracelular está también acumulado alrededor de la hifa de la endomicorrizas como punto de entrada a la célula hospedante y acumulados en los espacios intercelulares de la cubierta del hongo en las ectomicorrizas (Ashford et al., 1989; Smith & Smith 1990).
2.1.1 Tipos de interfase
Uno de los prerrequisitos para el movimiento de los nutrientes entre los simbiontes es el desarrollo de una interfase distintiva, la cual es modificada, estructural y funcionalmente, para promover la transferencia. En una asociación el desarrollo de la simbiosis es un proceso dinámico el cual puede resultar en la formación de diferentes tipos de interfases en diferentes tiempos y que la función de estas interfaces puede variar considerablemente.
Generalmente, son dos tipos de interfase que están involucradas en el transporte de nutrientes entre los simbiontes, denominados intercelular e intracelular. El desarrollo intercelular es ejemplificado por asociaciones ectomicorrizicas, en las cuales el contacto entre el hongo y el hospedante puede ser entre la envoltura del hongo y la capa externa de células de la raíces o entre las células corticales y la hifa de la red de “Harting” (o ambos). Solo en estados tardíos de desarrollo el hongo penetra las células de las células corticales de la raíz. En este ejemplo la interfase está compuesta de las membranas de los dos organismos separados por estas paredes, las cuales, con cualquier espacio intercelular, constituye una interfase en el apoplasto. La fase intercelular se presenta en muchas micorrizas vesiculo-arbusculares, como la hifa crece longitudinalmente en los espacios intercelulares de la corteza de la raíz.
En algunas asociaciones se ramifica la hifa intercelularmente, penetra la pared de la célula hospedante y crece entre ésta, formando la fase intracelular tales como arbúsculos, hifas o hasturio. En muchos de los casos es únicamente la pared, que es penetrada, la membrana plasmática de la célula hospedante mantiene intacta de tal manera que el microsimbionte se encuentra en el apoplasto intercelular. Las estructuras intercelulares y sus interfases con las células de la planta son generalmente consideradas las más importantes en el transporte de nutrientes que las interfases interceulares y esto es evidencia para esto en el caso de algunos parásitos formadores de haustorio. En algunas asociaciones de micorrizas VA la fase intercelular es reducida, la hifa pasa directamente de una célula cortical a otra, formando espirales o arbúsculos (Smith & Smith, 1990).
2.1.2 Localización de la simbiosis en el apoplasto
En las raíces, el hongo micorrízico coloniza el apoplasto cortical, inter o intracelularmente. De nuevo, conexiones plasmo desmátales entre las células influencian el balance entre el transporte simplástico y apoplástico y de esta manera el hongo se suple de los nutrientes. La endodermis forma una barrera para el movimiento apoplástico entre la estela y la corteza, y en las raíces de muchas especies una hipodermis suberizada puede prevenir el movimiento apoplástico entre la corteza y el suelo. Estas dos capas celulares efectivamente aíslan el apoplasto cortical, así que hay un control simplástico de la concentración de nutrientes del apoplasto cortical.
Tanto en raíces con ectomicorrizas y MVA la hipodermis bien desarrollada, el movimiento de nutrientes del suelo a las raíces estará restringido a una ruta simplástica a través de la hifa fungal y el paso a las células de la hipodermis. Los factores que afectan la pérdida de nutrientes minerales de la hifa fungal está determinada por el suministro de nutrientes al apoplasto cortical y también la toma por las células corticales. Recíprocamente un suministro de carbohidratos al hongo dependerá de la descarga a partir de las células corticales al apoplasto de las raíces. Las capas impermeables en los límites de la corteza (endodermis) pueden tener importancia en permitir un mayor control de las concentraciones apoplásticas en la corteza da ambas clases de raíz (Smith & Smith, 1990).
2.2 mecanismos de Transferencia
Los azúcares son importantes en la transferencia de carbohidratos, con la hidrólisis de la sacarosa. Otro recurso potencial de carbono incluye precursores de la pared, el cual puede igualmente requerir hidrólisis, y aniones orgánicos o componentes orgánicos de N.
Los nutrientes derivados del suelo que son transferidos del hongo a la planta varían en importancia en los diferentes tipos de asociación. Las Ectomicorrizas tienen mayor influencia sobre el nitrógeno (N) y un menor efecto sobre la nutrición en fósforo (P). En VAM la situación es inversa. La transferencia de N del hongo a la planta ha sido demostrada en Ectomicorrizas y es probable en asociaciones VAM. El N orgánico (como aminoácidos o amidas), más bien como NH4+, está probablemente involucrado. Tal transferencia puede resultar en el movimiento de carbono orgánico en contra del flujo de carbohidratos y afectaría el flujo neto de carbono reducido, dando una posible ruta para el movimiento del carbono durante la transferencia entre la planta. En ectomicorrizas se evidencia que el ortofosfato inorgánico es la mejor forma en la que el fósforo es transferido. En experimentos se ha demostrado la transferencia de otros nutrientes (S, Zn, Cu, Ca y Na) en varios tipos de micorrizas (Smith et al., 1994).
El movimiento de carbohidratos de la planta al hongo y el movimiento de nutrientes minerales a la planta ocurre al nivel de toda la planta. Asumiendo que la transferencia bidireccional ocurre en una simple interfase, ambos simbiontes tienen membranas plasmáticas funcionales capaces de tomar nutrientes del apoplasto. En las ectomicorrizas, la coexistencia de la actividad ATPasa en la membrana plasmática del hongo a la red sugiere que los dos sistemas trabajan cooperadamente en intercambio bidireccional (Smith, 1994).
2.3 toma de Macro y micronutrientes
Fósforo (P)
Los MVA pueden mejorar considerablemente el suministro de P para la planta hospedante mediante el uso de la capacidad absorbente de la extensa red de hifas externas asociadas a la raíz infectada. Las hifas de MVA, ligadas a la raíz, se extienden más allá de la zona de agotamiento de P que se desarrolla en los suelos deficientes en P, ya que la raíz absorbe iones fosfato más rápidamente de lo que se pueden difundir por el suelo para reabastecer el suministro a nivel de la superficie de la raíz. El movimiento de fosfatos que está mediado por las MVA ocurre en tres etapas: la absorción por la hifa endofita en el suelo, la translocación a la hifa dentro de la corteza radical mediante los puntos de entrada y la liberación hacia la planta. El hongo probablemente toma el fosfato del suelo, principalmente en la forma de iones ortofosfato de las existencias inestables.
Se cree que la translocación de este fosfato a la raíz ocurre principalmente mediante flujo citoplasmático de los gránulos polifosfatados en las vacuolas del hongo. Los MVA mejoran sustancialmente el crecimiento de las plantas a las cuales se les ha suministrado fuentes de fósforo relativamente insolubles, tales como harina de hueso, fosfato tricálcico y fosfato de hierro y aluminio. El P adicional fue tomado de la fracción de P soluble asociada con dichas fuentes fosfatadas o liberados por ellas en el suelo. Por consiguiente, las micorrizas aseguran una mejor utilización del fosfato disponible, en lugar de la movilización de la fracción insoluble (Saif, 1984).
El fosfato está presente en el suelo entres formas: (i) P inorgánico soluble en la solución del suelo, (ii) P inorgánico insoluble encontrado en estructuras cristalinas, y (iii) componentes orgánicos tales como Phytate. El fósforo en el suelo es relativamente inmóvil y se difunde lentamente a las raíces de la planta. Como resultado, en suelos con baja disponibilidad de P, las zonas de depleción se desarrollan alrededor de las raíces. En los suelos la cantidad de P que es disponible es poca y la cantidad es cerca del 1 al 5% del total del contenido de P (Cooper, 1984).
En general, el mayor efecto en el desarrollo por la infección con micorrizas es causado por el incremento en la absorción de P. La efectividad para la toma de P por el hongo está relacionada por: (i) Formación de polifosfatos en la hifa y, mantenimiento de concentraciones internas bajas de fosfato (Pi), (ii) el diámetro pequeño de la hifa proporciona un volumen relativamente grande en el suelo por unidad de superficie comparada con el área superficial de la raíz, y (iii) producción de ácido fosfatasa extracelular el cual cataliza el P liberado de los complejos orgánicos del suelo (Marschner & Dell 1994). Se calcula que la concentración de P en la parte externa de la hifa es alrededor de 0.3% de la materia seca comparada con las concentraciones del suelo "1 a 3 µM de ortofosfato (Pi). El P se absorbe como ortofosfato y es transferido a las hifas en forma de polifosfato, esta es la principal reserva de P en el hongo; los gránulos de polifosfatos se encuentran dentro de los arbúsculos jóvenes y en las vacuolas de las hifas. El modelo de transferencia de P a través de la interfase, generalmente involucra el flujo pasivo de Pi del hongo y activo de la absorción en la membrana plasmática de la célula de la planta. Cuando el hongo no se encuentra en simbiosis, la pérdida neta de Pi es poca, de esta manera las condiciones que promueven el flujo probablemente son importantes en la interfase hongo/raíz. El arbúsculo es un órgano del haustorio especializado capaz de gran actividad metabólica y particularmente está adaptado como un sitio de intercambio de nutrientes entre el hongo y la planta hospedante (Cooper, 1984).
La concentración intercelular de Pi en ambos, hongo y planta, afecta el transporte de Pi. Altas concentraciones de P en la interfase de la hifa pueden, con el tiempo, reducir la reabsorción, maximizando la pérdida del hongo. Contenidos bajos de fósforo en las raíces en relación al hongo (mantenidos por el metabolismo y/o translocación ente la planta) y el concomitante incremento en el activo transporte de Pi a la membrana plasmática de la planta, puede permitir el movimiento neto de Pi dentro de la planta. La transferencia de Pi al tejido hospedante en ectomicorrizas es cerca de 10-20% del Pi absorbido por la hifa (Marschner & Dell, 1994; Smith, 1994).
2.3.2 Nitrógeno
Las investigaciones en toma de nitrógeno en plantas con VAM han sido en leguminosas. Cuando VAM mejora la nutrición de P en la planta hospedante, esto puede corresponder a un incremento en la nodulación, fijación de N2 y desarrollo. En vista de los altos requerimientos de P para la nodulación, muchas especies de leguminosas que se desarrollan en suelos bajos de P son altamente dependientes de la infección VAM. Sin embargo, la simbiosis impone una gran competencia por los fotosintatos, usualmente a expensas del crecimiento de las raíces. De acuerdo a los efectos benéficos en la fijación con N2 son uno u otro confinados a suelos con bajo P (Marschner & Dell, 1994).
Estudios con radioisótopos revelan que el micelio externo del hongo puede utilizar el N inorgánico del suelo eficientemente y es transportado 10-30 cm a través del suelo. Así, las plantas micorrizadas tienen acceso a formas de N que no son disponibles para plantas sin esta asociación. Bago et al. (1996) mostraron que Glomus intraradices es capaz de tomar NO3- y translocarlo a la planta. En muchos suelos agrícolas el ión NO3- es la forma predominante de N; aunque este ión es altamente móvil, cuando los suelos están restringidos por el contenido de agua, su movilidad se reduce drásticamente. En estas condiciones, el papel de las micorrizas en el transporte de NO3- a las raíces puede ser significativo. Estudios realizados en maiz (Zea mays) infectados con G. intraradices, evidencia que el transporte de la hifa a la planta fue del 33%, aunque puede variar con la compatibilidad funcional del hongo. Johansen et al., (1996) indican que ambas formas de nitrógeno NO3- y NH4+ son asimiladas en el micelio del hongo. Esta forma de incremento de toma de N puede estimular las enzimas involucradas en la asimilación de nitrógeno de la planta hospedante. Estos datos sugieren una relación positiva entre la contribución de N por la hifa y el estado metabólico/nutricional de la planta. Estos cambios pueden ayudar a la planta para resistir condiciones secas del suelo (Subramanian & Charest 1999).
Micronutrientes
La movilidad de Cu, Zn, Mn y Fe en suelos es baja. Como resultado, la toma de estos nutrientes por las raíces es limitada. Cuando no hay adición de estos micronutrientes al suelo, la disponibilidad es aún más baja y se forman zonas de depleción alrededor de las raíces. Las plantas micorrizadas pueden tomar más nutrientes metales vía hifa extraradical, la cual provee grandes áreas de superficie (Jakobsen et al., 1992). De acuerdo a Bürket y Robson (1994) la toma de Zn está influenciada por la distribución y longitud de la hifa en tres especies de VAM en el suelo. La alta densidad de la hifa en el suelo, gran superficie de absorción, corta distancia de los metales para difundirse, y la efectividad de plantas micorrrizadas pueden absorber estos nutrientes metales poco móviles.
Liu et al., (2000) encontraron que la contribución en la toma de Zn, Cu, Mn y Fe en maíz está significativamente influenciada por los niveles de P y micronutrientes en el suelo. Su efecto es positivo o negativo dependiendo no sólo del P sino también de los niveles de los micronutrientes en mención. La hifa extraradical puede absorber y transportar Cu y Zn a su planta hospedante. El efecto benéfico de la inoculación con micorriza para la toma de Cu y Zn se elimina con la adición de estos micronutrientes al suelo. Bajo condiciones de Cu y Zn abundante, las raíces del maíz parecen depender menos de la micorriza. El efecto de P sobre la toma de Zn tiene dos componentes. Por un lado, las raíces de las plantas crecidas en bajo contenido de P poseen más hifas y, sin embargo, puede potencialmente absorber más Cu y Zn.
Koothari et al., (1991) reportaron en plantas de maíz crecido en suelos calcáreos, la contribución de la hifa de G. Mosseae a la toma total en un rango entre 16 y 25% de Zn y 13 - 20% de P.
2.4 Importancia de la transferencia de nutrientes en la productividad de la planta
Los hongos de MVA son abundantes y de suma importancia ecológica en el trópico. Extensas áreas de suelos tropicales tales como el Cerrado de Brasil y los Llanos Orientales de Colombia, son deficientes en P o inmovilizan fertilizantes de P. Estas tierras agrícolas marginales podrían ser productivas si se desarrollan y adicionaran al suelo hongos de micorrizas con la capacidad de utilizar cantidades extremadamente pequeñas de fertilizantes. Como fuentes de P se podrían adicionar las rocas fosfatadas de bajo costo.
Se ha demostrado que algunos hongos de micorrizas utilizan la roca fosfatada mucho mejor que otras fuentes y pueden mejorar considerablemente el crecimiento de las planas que se desarrollan en suelos pobres fertilizados con este material. Una alternativa viable al uso de fertilizantes de alto costo en suelos agrícolas pobres que han sido talados y quemados, puede ser el uso de hongos de MVA efectivos en el trópico.
La transferencia entre los simbiontes está basada en el efecto potencial de la infección sobre la producción de la planta, o su supervivencia. Cuando el crecimiento y reproducción de la planta es limitada por la tasa de adquisición de nutrientes y cuando la infección micorrizica incrementa esto (por procesos de toma y transferencia), es claro que la infección es de importancia y que el resultado neto de la transferencia bidireccional en un parámetro medible en la biomasa de la planta. Sin embargo, la infección por la micorriza no siempre incrementa el crecimiento de la planta o su reproducción (Smith, 1994).
| Concentración en materia seca de raíces | |||||||
| mg g-1 |
|||||||
| Enviado por: | Germán Colmenares |
| Idioma: | castellano |
| País: | Colombia |