Biología, Botánica, Genética y Zoología
Mecanismos de defensa en plantas
1. INTRODUCCIÓN
Los mecanismos de defensa (o estrategias de afrontamiento) son procesos psicológicos automáticos que protegen al individuo frente a la ansiedad y las amenazas externas o internas, y en general individuo, en este caso la planta suele ser ajeno a estos procesos. En la naturaleza las plantas están expuestas a un gran número de organismos patógenos como insectos, hongos, bacterias y virus. Es entonces sorprendente constatar que, a pesar de la inamovilidad y la ausencia de un sistema inmune, la muerte de plantas por enfermedad termine siendo una situación excepcional. Esto hace pensar que los mecanismos de defensa de las células vegetales, tanto aquellos constitutivos como aquellos inducidos por la presencia del patógeno, son muy efectivos para detener o contrarrestar una infección. Además se ha comprobado que las plantas son capaces de reaccionar ante estímulos externos, como puede ser la intensidad de luz, la cantidad de agua o la presencia de algunas sustancias en los alrededores. Las plantas poseen un patrón de conducta lineal, según el cual actúan de forma directa ante cualquier estímulo. La agresión que sufre la planta desencadena una serie de reacciones químicas por las que se liberan unas sustancias al aire. Para ello, es imprescindible la presencia de una serie de componentes que la propia planta sintetiza, como es el caso de las proteínas. Además han tenido que sufrir adaptaciones a estrés abiótico y elaborar estrategias hábiles para la defensa y hacer frente a las agresiones tanto de tipo biótico (insectos, hongos, virus), como de tipo abiótico (agua, calor, oxido nítrico).
2.1. Las plantas tienen una doble línea defensiva
Las plantas están expuestas a muchos agentes patógenos diferentes en el entorno. Sin embargo, sólo algunos pueden afectarlas. Si un agente patógeno particular es incapaz de atacar con éxito a una planta, se dice que ésta es resistente a aquél; en otras palabras, que no puede hospedar al patógeno.
Aunque en la naturaleza este tipo de inmunidad detiene a casi todo parásito, ha sido objeto de poca investigación. Algunos científicos han desvelado los componentes moleculares de esa primera línea defensiva. En sus descubrimientos, deducen paralelismos entre los sistemas inmunológicos vegetales y animales.
Los investigadores han constatado que el gen conocido como PEN es un componente importante del mecanismo de resistencia en la primera línea defensiva. Aislaron mutaciones de plantas Arabidopsis parcialmente susceptibles a algunos mohos, como el de la especie Blumeria graminis. Si estos genes son defectuosos o si falta la proteína que codifica en las células de la planta, el hongo puede invadir la epidermis foliar. Por eso, los científicos examinaron cuál es exactamente la función de la proteína PEN2 en la defensa contra los agentes patógenos.
La planta idopsis thaliana se defiende contra un parásito invasor. A la derecha, con el gen PEN2 desconectado.
La PEN2 es una enzima localizada en la membrana de los llamados peroxisomas. Éstos son compartimientos celulares separados, en los que a menudo tienen lugar reacciones metabólicas que serían peligrosas para el organismo si ocurrieran en otro lugar. Si un hongo trata de invadir una célula vegetal, los peroxisomas son conducidos al sitio de entrada mediante la PEN2. Una o más moléculas de azúcar pueden separarse de otro componente celular a través de la actividad enzimática de la PEN2, una glicosil hidrolasa. La sustancia liberada parece tener un efecto fungicida, que mata al patógeno.
Sin embargo, en ausencia de PEN2, la planta no está completamente indefensa contra los hongos atacantes. Existe una segunda línea de defensa que es una medida drástica: la célula muere conjuntamente con su agresor, protegiendo al tejido circundante de la infección.
En esta línea defensiva suicida, proteínas muy diferentes desempeñan un papel crucial, particularmente las conocidas como EDS1, PAD4 y SAG101. Sólo si este segundo mecanismo también falla, puede producirse una colonización plena por Blumeria graminis.
2. 2. PROTEINAS INVOLUCRADAS EN EL MECANISMO DE DEFENSA DE LAS PLANTAS
Las plantas, careciendo de un sistema de defensa basado en anticuerpos similar al que existe en los animales, basan su protección en características físicas y en una serie de componentes que la propia planta sintetiza. Dentro de esos compuestos, las proteínas constituyen una de las principales fuentes de defensa, no sólo por su elevada especificidad y eficiencia, sino porque además algunas de ellas son altamente reguladoras, respondiendo su síntesis al ataque de los depredadores (insectos) o de los patógenos. Estas proteínas representan una interesante alternativa para producir plantas con mejores características de resistencia, ya que a través de mecanismos de fitomejoramiento o bien introduciendo el gen a plantas sensibles, por medio de la ingeniería genética. El conocimiento de sus mecanismos de acción representa una forma importante para aprender a combatir plagas y enfermedades de las plantas, sin tener que utilizar compuestos altamente contaminantes, como son los insecticidas comerciales. El constante ataque de insectos y de patógenos sobre las plantas y granos almacenados, produce pérdidas muy elevadas para los agricultores, quienes han tenido que recurrir al uso intensivo de pesticidas. Sin embargo, su utilización presenta grandes problemas, ya que al no ser específicos contra una clase particular de organismos, y dado que su toxicidad abarca una amplia variedad de organismos, han generado daños considerables tanto a la ecología como a los organismos superiores. Además al llegar las lluvias, éstas arrastran a los insecticidas, produciendo serias contaminaciones de los mantos freáticos. Es por ello que se estudian mecanismos que permitan generar plantas con mayor resistencia al ataque de insectos y de patógenos, tales como los hongos, virus y nematodos, buscando mecanismos naturales que tengan una elevada selectividad y que además, en las condiciones en las que el producto se ingiere, no presenten toxicidad contra los humanos o contra los animales. Las plantas han generado con la evolución una serie de sistemas de defensa contra el ataque de insectos y patógenos basados en barreras estructurales o químicas o generando una respuesta metabólica activa. La resistencia de las plantas está frecuentemente dividida en defensa constitutiva, expresada como una característica normal del desarrollo de la planta, o defensa inducible, la cual se activa al contacto con un organismo invasor. Este último requiere un sistema de vigilancia, el cual permita el reconocimiento de la amenaza, generando un sistema de transducción de señales y una ruta de respuesta, usualmente regulada a nivel transcripcional por medio de la expresión de genes relacionados con la defensa. Existe un número elevado de proteínas que han sido descritas como parte de los mecanismos de defensa de las plantas, las cuales, sólo actuarían sobre los depredadores que ataquen a la planta o a los granos, no produciendo ningún efecto sobre otros organismos. Por esta razón, es importante considerar que estos mecanismos no deben ocasionar daño cuando el hombre o los animales los utilizan como alimento. Esto puede ocurrir debido a su estricta selectividad que le permita reconocer como blanco solamente al insecto o patógeno correspondiente.
Interacciones de plantas con parásitos y depredadores
Existe una enorme variedad de agentes bióticos con el potencial de depredar o parasitar tejidos de plantas, entre los que se incluyen invertebrados, especialmente insectos, nematodos y microorganismos tales como hongos, bacterias, micoplasmas y virus. Las relaciones entre estos agentes y la planta huésped también varían desde transitorias en tejidos expuestos, hasta asociaciones íntimas de largo tiempo con un alto grado de dependencia y especificidad. Muchos hongos parasíticos de plantas, algunos nematodos e insectos y todos los virus son completamente dependientes de células vegetales vivas para completar su ciclo de replicación.
Tipos de defensa de plantas
La resistencia de las plantas a plagas y patógenos es compleja y basada en la acción combinada de varios factores.
En la resistencia a invertebrados, tal como sucede con los insectos, se incluyen factores que modifican el comportamiento del animal sobre la planta huésped, así como los que afectan la calidad de la planta como fuente de alimentación. Así, Los componentes estructurales o químicos, pueden actuar para repeler o disuadir al insecto de la planta seleccionada como fuente de alimento. Si tales defensas fallan, entonces, otros componentes tales como los antinutricionales, pueden asegurar que la planta sea un huésped pobre en cuanto a valor nutricional, limitando así el grado de daño. La respuesta activa al ataque incluye efectos sistémico tales como la síntesis de inhibidores de proteasas en tejidos lejanos al sitio inicial de daño.
Los factores constitutivos, tanto anatómicos como químico tales como: cutícula, pared celular e inhibidores enzimáticos, son utilizados para prevenir la colonización de tejidos de la planta, por la mayoría de los microorganismos no patogénicos. Si ocurriera una penetración por parte del microorganismo, entonces los sistemas de defensa inducibles serían activados. Esto incluye una rápida generación de especies activas de oxígeno, cambios en los polímeros de pared celular, síntesis de compuestos de bajo peso molecular como las fitoalexinas, producción de proteínas relacionadas con la defensa, y muerte celular hipersensitiva. Colectivamente estos sistemas primero inhiben y después sellan al colonizador potencial. En términos genéticos es usual distinguir entre genes de reconocimiento, los cuales codifican proteínas receptoras o asociadas con rutas de transducción de señales, y genes de respuestas, los cuales codifican proteínas que actúan como factores de defensa o como enzimas biosintéticas en rutas que dirigen a la producción de compuestos de defensa. Las proteínas de las plantas además tienen papeles claves en muchos aspectos de la defensa, tanto como factores constitutivos de resistencia, como formando parte de la compleja cascada de respuesta de resistencia.
Proteínas protectoras, propiedades generales
Las proteínas protectoras pueden ser divididas en dos grandes grupos basados en sus patrones de expresión, aunque hay una considerable sobreposición entre ellas. El primer grupo de proteínas puede ser descrito como constitutivo o tejido-específico, y su expresión no está relacionada a la infección o daño, aunque éstas pueden estar restringidos a órganos, tejidos o tipos de células específicos. Éstos están particularmente diseminados en semillas y otros tejidos de almacenamiento donde su presencia podría estar confiriendo resistencia a un daño o infección.
Las proteínas protectoras constitutivas más ampliamente estudiadas son los inhibidores de enzimas hidrolíticas, principalmente de proteasas y amilasas. Estos inhibidores, los cuales están frecuentemente presentes en altas cantidades, actúan como proteínas de reserva.
Los inhibidores de enzimas de plantas están clasificados en familias, basados en la secuencia de aminoácidos, estructura y sus especificidades. En el caso de los inhibidores de proteasas, los mejor estudiados han sido los de serín proteasas como tripsina, quimotripsina y subtilisina. Sin embargo, también se han caracterizado inhibidores de otra clase mecanística, como carboxipeptidasas y cisteín proteasas, así como los inhibidores de (alfa)amilasa, aunque estos últimos pueden ser altamente específicos para enzimas de una particular fuente(por ejemplo, algunos tipos de insectos, bacterias o hongos).Pero algunos inhibidores podrían tener otras actividades como puede inducirse por su semejanza con secuencias de aminoácidos de proteínas conocidas.
Las semillas son ricas fuentes de muchas otras proteínas que tienen funciones protectoras. Por ejemplo: la cebada, que contiene tioninas, endoquitinasas, proteínas que inactivan ribosomas(RIP), B-glucanasas, LTP, lectinas, peroxidasa, y proteínas tipo taumanina.
El segundo gran grupo de proteínas protectoras exhiben patrones de expresión más altamente regulados, siendo inducidos a respuesta a infección o daño. Éstas, incluyen las proteínas relacionadas con la patogénesis(PR), que además están presentes en el tabaco. Se localizan en las vacuolas más que en el apoplasto, y están, involucradas con la defensa de plantas, como las quitinasas y proteínas antifúngicas.
Otras proteínas relacionadas con la defensa inducida por infeccion o daño incluyen los inhibidores de proteasas PI y PII de papa. Estos inhibidores de tripsina y quimotripsina, no sólo son inducidos por daño mecánico en hojas, sino también son expresados en tubérculo y flores.
Dos de los principales grupos de las proteínas PR inducidas por infección en tabaco por el virus del mosaico del tabaco, son las B-1,3-gluacanasa y endoquitinasa. Ya que los b-glucanos y la quitina están presentes en la pared celular de muchos hongos patógenos. Las endoquitinasas de plantas frecuentemente muestran actividad de lisozima, y algunas lisozimas de endoquitinasa; ambas actividades requieren de la hidrólisis de enlaces B-1,4 entre homopolímeros de N-acetilmurámico y N-acetilglucosamina para lisozima. Los lisozimas inhiben el crecimiento bacteriano por hidrólisis de peptidoglicanos de la pared celular, por lo que juegan un papel en la defensa contra bacterias patógenas.
Resistencia a patógenos microbianos
Durante la colonización de tejidos, los microorganismos patógenos de plantas, especialmente los patógenos necrotróficos, despliegan enzimas hidrolíticas extracelulares contra polímeros de pared celular. Tales enzimas, incluyen pectato liasas y pologalacturonasas, las cuales han sido implicadas como factores de patogenicidad de bacterias y hongos patogénicos de plantas.
Proteínas que inhiben poligalacturonasa han sido detectadas en una amplia variedad de plantas dicotiledóneas, y en frutos tales como tomate, manzana y pera. Las poligalacturonasas pueden actuar como factores potenciales de virulencia, por la liberación en presencia de PGIP de moléculas señal que activan la ruta de defensa de la planta.
Inhibidores de proteasas
En las plantas los polipéptidos y proteínas proteolíticas están generalmente concentradas en tejidos de almacenamiento, como semillas y tubérculos. Su abundancia en estas familias no es única, ya que se han encontrado en raíces y hojas en géneros de otras familias.
Los inhibidores de proteasas son elementos importantes de la respuesta de defensa de la planta a la degradación. El papel de los inhibidores en la regulación de las actividades proteolíticas es muy importante por la protección que proporcionan a tejidos y fluidos de la degradación no deseada. Los inhibidores de proteasas encontrados en plantas, generalmente contienen altos porcentajes de cisteína formando puentes disulfuro, lo cual los hace resistentes a la desnaturalización por calor y ph extremos. La producción de estos inhibidores está altamente regulada por una ruta de transducción de señales que es iniciada por el ataque del patógeno y transducida como una respuesta de daño.
Los inhibidores de proteasas se acumulan durante la maduración de la semilla, y se ha argumentado que la maduración de semillas es un indicador de su función de defensa.
Con el ataque de herbívoros se inicia la respuesta de defensa de la planta que incluye la inducción de la expresión de genes de inhibidores de proteasas que pueden estar ligados directamente con la resistencia al ataque del insecto. La inducción de expresión de genes de inhibidores de proteasas ocurre tanto en células del sitio del daño, como en lugares distantes a ese sitio. La respuesta de defensa puede incluir la expresión de múltiples inhibidores de proteasas que pueden inhibir un amplio número de proteasas digestivas del herbívoro.
Respecto al daño mecánico, los fragmentos de oligosacáridos péctidos que son liberados de la pared celular de la planta, y oligómeros de quitosana derivados de la pared celular de hongos, se asume que actúan como inductores extracelulares de la ruta de señales que dirige a la expresión de genes de inhibidores de proteasas. Los polisacáridos inductores no parecen ser inmóviles en la planta, por lo que es probable que estas moléculas únicamente induzcan en forma local la expresión de genes de inhibidores de proteasas. La magnitud de respuesta de defensa es mucho mayor en plantas atacadas por insectos que en plantas dañadas mecánicamente.
3. Tipos de agresiones: Estrés biótico y abiótico.
Los vegetales son hipersensibles y, por su imposibilidad de huir, han elaborado estrategias asombrosamente hábiles de defensa activa y pasiva para hacer frente al estrés al que están sometidos. Explican los científicos que las plantas están expuestas a dos tipos de agresiones: las que proceden de otros seres vivos, como el hombre, los animales, los hongos o los virus, que se denomina estrés biótico. La otra procede del ambiente, como la sequía, el calor o el viento, que se le conoce por estrés abiótico.
3.1. Estrés biótico en vegetales:
Estrés biótico se denomina al conjunto de agresiones que padecen las plantas y que viene producido por otros seres vivos, como insectos, hongos, virus, agentes patógenos, plagas, etcétera. Este estrés provoca en la planta la pérdida de rendimiento.
A continuación se detalla y se explica los tipos de agresores más importantes:
% Mamíferos herbívoros:
Los herbívoros ejercen una gran presión sobre las plantas, por ejemplo, en el bosque mediterráneo, cada vez aumenta más el número de herbívoros debido a la caza, la explotación ganadera, etc. Los herbívoros pueden limitar la regeneración forestal, ya que consumen semillas y plántulas jóvenes. Sirva como ejemplo el caso del pino albar en Sierra Nevada, del cual los depredadores consumen casi el 99% de sus semillas, lo cual constituye una dificultad mayor para que la semilla del pino pueda germinar. En ocasiones, las plantas usan como mecanismo de defensa la asociación con otras plantas más resistentes que ellas, este es el caso de algunas plantas del bosque, que crecen en zonas de matorral el cual los protege del pisoteo y la depredación de la herbivoría al hacer de barrera entre dichos herbívoros y la planta.
%Insectos y plagas:
Los insectos y las plagas producen en las plantas un daño mecánico sobre todo provocado por la acción de insectos fitófagos. Las plantas reaccionan al ataque por plagas activando una batería de genes de defensa en el tejido dañado y también en zonas distales no dañadas en la planta herida. La planta transporta una enzima hacia el lugar de infección. Puesto que tiene un efecto fungicida, allí mata al patógeno, si este sistema falla, entra en acción la segunda defensa de la célula, que consiste en provocarse la muerte y, de ese modo, producírsela también al patógeno, de esta forma consigue poner a salvo al resto de la planta del contagio de dicho agente patógeno.
Las plantas interaccionan con los insectos depredadores, para percibir la presencia de éstos, las plantas detectan las secreciones orales de éstos. Entre las defensas vegetales directas inducidas por los herbívoros destaca la producción de metabolitos secundarios toxicos o repelentes y de moléculas volátiles. Estas desempeñan también un importante papel en le defensa indirecta de la planta. Algunos compuestos volátiles suelen ser comunes a muchas especies, como alcoholes, aldehídos, ésteres y terpenoides. Otros compuestos son, por el contrario específicos de cada especie. Los hay que sirven para atraer depredadores y parásitos que destruyan a los herbívoros agresores, es decir, utilizan la ayuda de otros artrópodos. En otros casos, la porción de ácido graso de ciertos compuestos, como la volicitina, sufre una modificación en el intestino de los insectos, que concluye en la emisión de compuestos volátiles los cuales atraen parásitos del insecto, por lo que ayudan a la planta a deshacerse del insecto no deseado. Esta liberación de sustancias volátiles puede reducir el ataque de la herbivoría hasta en un 90%. Pese a esto, y como resultado de la coevolución, algunos insectos herbívoros se han adaptado a las defensas de las plantas, se puede observar que los coleópteros y los lepidópteros hacen frenta a la presencia de los inhibidores de proteasas mediante la síntesis de proteasas insensibles a la inhibición, esto es una capacidad propia de cada especie.
%Agentes patógenos:
En la naturaleza las plantas están expuestas a un gran número de organismos patógenos como insectos, hongos, bacterias y virus. Sin embargo, sólo algunos pueden afectarles. Si un agente patógeno particular es incapaz de atacar con éxito a una planta, se dice que ésta es resistente a aquél; en otras palabras, que no puede hospedar al patógeno. Pese a esta gran variedad de estresantes bióticos y ante la ausencia de sistema inmune y movilidad de las plantas es difícil que una planta muera por enfermedad. Esto hace vigente que los mecanismos de defensa de las células vegetales, tanto aquellos constitutivos como aquellos inducidos por la presencia del patógeno, son muy efectivos para detener o contrarrestar una infección.
Las plantas presentan barreras estructurales que intentan impedir la entrada de patógenos. Entre ellas se encuentran la pared celular, lignina, cutícula, y las aperturas naturales de las plantas (estomas, lenticelas, etc.). Para superar estas barreras, los patógenos necesitan de enzimas hidrolíticos que destruyan estas barreras estructurales, así algunos producen peptinasa que afecta a la pared celular de las plantas, otros cutinasa que altera la cutícula, etc. Además de estas barreras, las plantas acumulan metabolitos secundarios, que en muchos casos poseen propiedades anta-microbianas, y que van a actuar en el lugar de penetración del patógeno.
Tras el ataque de los patógenos a la planta, ésta pone en marcha mecanismos de respuesta local como son la lignificación, la formación de papilas, el enriquecimiento de la pared celular en glucoproteínas ricas en hidroxiprolina, o la oclusión vascular. Junto al desarrollo de estructuras locales para bloquear la entrada de los patógenos, también se acumulan sustancias alrededor del lugar de la infección, conocidas como fitoalexinas, que incluyen una gran cantidad de productos naturales, que se producen en mayor cantidad tras la entrada de un patógeno y poseen capacidad antimicrobiana.
Además de la defensa local, la resistencia de las plantas puede extenderse hacia regiones totalmente intactas de la misma que no han sido lesionadas por el patógeno. Esta resistencia se ha denominado Sistémica Inducida o Adquirida. La respuesta tiene una duración de semanas y no es específica, ya que actúa de forma semejante para una amplia variedad de patógenos, o incluso tras lesiones mecánicas no causadas por patógenos invasores (la producción de pequeñas lesiones a las plantas ha sido utilizada por agricultores desde antiguo para inducir resistencias en las mismas frente a la invasión de diversos patógenos).
El mecanismo a través del cual las plantas reconocen a los patógenos no es del todo conocido, pero parece que componentes que se liberan durante la interacción planta-patógeno, conocidos como inductores, van a ser reconocidos por receptores presentes en la planta. La interacción con estos receptores llevará a la activación de genes de resistencia sistémica y a la síntesis de las proteínas antes comentadas denominadas proteínas de resistencia sistémica (PRS).
Reacción hipersensibilidad provocada por la infección de plantas de tabaco con el virus del mosaico del tabaco (TMV). Se muestran las lesiones necróticas que se desarrollan en los sitios de ingreso del patógeno.
3.2. Estrés abiótico en vegetales:
La evolución transcurrida durante millones de años, ha condicionado su gran diversidad que resulta de la adaptación frente a distintas condiciones ambientales.
La diversidad de especies vegetales existentes, resultante de una evolución continua en el tiempo, ha debido implicar el desarrollo de mecanismos fisiológicos y morfológicos que han hecho posible la adaptación de las plantas a los hábitats más diversos y más extremos del planeta. Forma y función han estado siempre relacionados y se complementan para optimizar la vida en ambientes diferentes, ya sea para dotar de tolerancia frente a rangos extremos de temperatura o donde existen limitaciones de disponibilidad de agua, ambientes permanentemente inundados, en suelos contaminados por el hombre o en sustratos por naturaleza adversos, las plantas se ajustan para su evolución a las diferentes condiciones de estrés del medio ambiente. Estas son algunas de las diferentes estrategias adaptativas:
TOLERANCIA A LA SALINIDAD Y A ELEMENTOS PESADOS
Las plantas pueden ajustar su metabolismo a diferentes condiciones negativas ya sea de extrema salinidad, en ambientes contaminados con elementos pesados o en presencia de residuos industriales. Muchas otras especies pueden tolerar la sequía y salinidad típica de desiertos y, alternativamente, un conjunto denominado "manglares" son plantas que pueden adaptarse y crecer aun sumergidas sus raíces en agua marina conteniendo 31 gramos de sal por litro aprox. Además, existen las denominadas especies "acumuladoras" que poseen la propiedad de almacenar en cantidades extremas uno o más elementos pesados y tóxicos para las plantas presentes en el suelo como Uranio, Cadmio, Plomo, Arsénico, Zinc, Cobre, Fierro, Níquel, Selenio, etc.
Las plantas acumuladoras pueden cumplir con un rol esencial vinculado a descontaminación de suelos al incorporar, acumular, metabolizar e inactivar varios de dichos elementos. Este proceso se llama "fitorremediación".
Existen varios mecanismos de tolerancia de las plantas frente a un exceso de iones y metales pesados en el ambiente. Entre ellos, junto con lograr un impedimento de ingreso al citosol, existe fijación de metales en las paredes celulares, formación de complejos inorgánicos, inmovilización por proteínas específicas (fitoquelatinas), unión e inactivación con ácidos carboxilicos, (uniones a malato, citrato, oxalato) y fijación a varios compuestos orgánicos a acumularse en la vacuola.
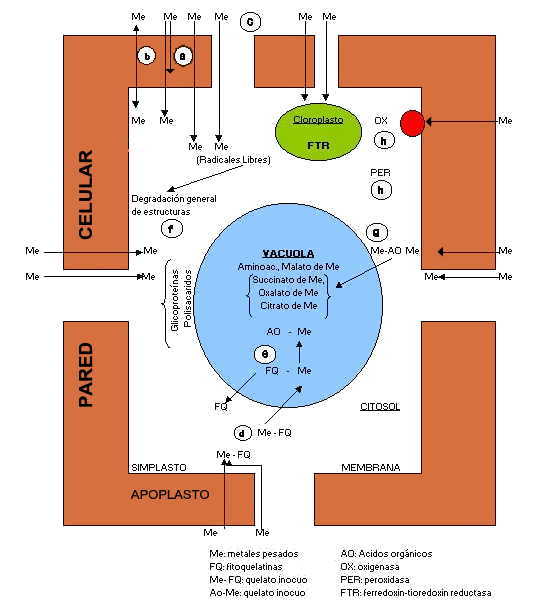
Mecanismos de Transporte, Detoxificación y Resistencia de las Células Vegetales frente a Metales Pesados.
(a) Inmovilización de metales en la pared celular y membrana plasmática (plasmalema); por acción de pectinas, proteínas, aminoácidos y otros componentes. (b) Transporte activo de metales (re-exportación) por carriers específicos a nivel de membrana. (c) Ingreso por pared (apoplasto) y membrana (simplasto) con posible formación de radicales libres en citosol. (d) Ingreso y fijación de metales pesados a fitoquelatinas (FQ) en el citosol formando compuestos quelados inocuos / fijación a polisacáridos y glicoproteinas en citosol / ingreso a vacuola. (e) Conversión Ingreso de compuestos complejados FQ a AO (malato, succinato, citrato, etc.) y compartimentación en vacuola. (f) Fijación de metales pesados a glicoproteinas y polisacáridos en el citosol / ingreso a vacuola. (g) Fijación de metales pesados a ácidos carboxilicos/orgánicos (AO) directamente en el citosol; ingreso a vacuola. (h) Acción de sistemas de defensa antioxidante en pared/membrana (OX=oxigenasa); citosol (PER=peroxidasas) y en cloroplastidios (FTR=ferredoxin - thioredoxin reductasa).
Los diferentes posibles daños que ocurren al no funcionar los mecanismos protectivos pueden ser: destrucción de ADN, de proteinas, membranas, reducción de glutationa, degradación de tilacoildes en cloroplastos, inhibición de enzimas del ciclo de Calvin y daños al sistema pigmentario II (PSII) y células vecinas.
Otra estrategia visible en algunas especies frente a la salinidad es la conversión paulatina de plantas C3 al metabolismo propio de las CAM ( ej. Cactus) implicando la síntesis de-novo de la enzima PEP-carboxilasa al cabo de unos 4-5 días de sometimiento a este tipo de estrés. La estrategia consiste en que a través de la presencia de la PEP-carboxilasa se permite la fijación de CO2 de noche, periodo en que los estomas de las plantas CAM se encuentran abiertos. Dichos mecanismos están referidos a reducir la pérdida de agua por transpiración, aumentando la tolerancia a sales y sequía frente a estas condiciones.
TOLERANCIA A LA SEQUIA Y ESTRES HIDRICO; EFECTOS DE TEMPERATURA VS ECONOMIA DEL AGUA EN ZONAS DESERTICAS.
AGUA. Tolerancia y adaptación también se aplica a aspectos de sequía en las plantas efecto que puede ser agravado al potenciar otros daños causados por altas temperaturas del ambiente. El concepto de sequía e implicancias sobre la marchitez, no debe vincularse sólo respecto a un contenido definido de agua en un suelo sino más bien a la disponibilidad de agua existente en el sustrato que la planta puede efectivamente usar .El agua ingresa a la célula vegetal en términos físicos de "energía libre". Ello implica difusión desde ambientes con mayor cantidad de moléculas de agua hacia donde se encuentra menos agua disponible. El rango de disponibilidad del agua que existe en el suelo (y el agua que puede difundir a las raíces) va desde niveles de aprox. -0.3 Atm. a -15 Atm. El agua retenida por el suelo a tensiones mayores no es transportada a las raíces y no puede ser usada por las plantas. Ambos rangos límites, que definen el agua disponible y la vida de la planta, corresponden a "Capacidad de Campo y Punto de Marchitez Permanente", respectivamente. A tensión de -15 Atm. la planta se marchita irreversiblemente aunque se someta inmediatamente a ofertas de agua óptimas para su recuperación. El problema del constante suministro del agua de las plantas a la atmósfera denominado transpiración.
TEMPERATURA. Las altas temperaturas a las cuales están expuestas las plantas son una limitante para que las hojas y tallos puedan mantener la estructuración de sus proteínas y enzimas requeridas para mantener el crecimiento. La necesidad de realizar reacciones químicas, metabolismo y construcción de estructuras celulares, implica estabilidad de mecanismos bioquímicos por lo cual deben ser impedidos registros de temperatura en hojas y tallos superiores a 55ºC. Condiciones como las anteriores por periodos mayores a 15 min. implican la muerte de las plantas. La gran estrategia evolutiva común de funcionamiento en plantas terrestres corresponde a desarrollar una mayor tasa transpiratoria cuando el agua no es limitante. La transpiración implica pérdida a la atmósfera de agua líquida en forma de vapor; el calor de evaporación cedido por las hojas reduce permanentemente su temperatura. Junto con la liberación de calor, existen otros procesos acompañantes y de apoyo para la adaptación al problema de economía del agua, que se citan más abajo:
-
Las plantas conforman productos que implican apoyo osmótico. Se ha visto que la porción apical y regiones de las hojas más expuestas a la luz y temperatura, sintetizan compuestos que estabilizan su condición y favorecen especialmente la retención de agua como defensa a la marchitez por sequía.
-
Las plantas pueden regular mecanismos temporales de cierre estomatal; producir cambios de posición y expresar caída anticipada de hojas. A través de la apertura estomática se regula simultáneamente la salida de agua y el ingreso de CO2 para el proceso de fotosíntesis. La gran mayoría de las plantas abren sus estomas de día y los cierran de. De acuerdo a la estrategia adaptativa, tal como ocurre en varias especies insertas en el grupo CAM, éstas por contrario abren sus estomas de noche, cerrándolos de día. La respuesta adaptativa de plantas CAM para fijar CO2 es activar otra enzima capaz de tomar el CO2 del ambiente. Mediante la acción de la PEP-carboxilasa presente en estas especies es posible fijar el CO2 en forma de malato (molécula de 4 carbonos) y confinar estos carbohidratos en la vacuola de las células. El malato formado de noche se transforma durante el día en piruvato (molécula de 3 carbonos) y CO2 el cual es captado por la enzima Rubisco que da inicio al ciclo de Calvin a estoma cerrado. La importancia adaptativa de este mecanismo de las plantas CAM reside en el hecho que éstas proceden de noche con máxima economía de agua.
La economía hídrica en estas plantas implica también variaciones drásticas de su morfología. Por otro lado, junto a un escaso número de estomas, la falta de hojas reduce la superficie transpiratoria y la posición vertical favorece una menor incidencia de luz. La regulación de la temperatura interna se realiza por difusión continuada del agua en su proyección horizontal. Durante el día se calienta más el agua celular de las células más externas, el agua caliente difunde hacia al interior, y ésta de menor temperatura se mueve hacia la periferia, enfriando. Durante la noche se produce el fenómeno inverso. También las espinas, se consideran morfológicamente relevantes en la economía hídrica. En muchas zonas desérticas, en localizaciones más cercanas a la costa y con alta humedad ambiental proveniente del mar, las espinas favorecen la condensación de la humedad durante la noche. Las gotas fluyen por gravedad por el tallo a la superficie arenosa del suelo donde pueden ser captadas por las raíces superficiales favoreciendo la economía del agua.
Otra estrategia es tener hoja ancha y no suculentas, consiste en acelerar la senescencia de sus hojas, y a través de su eliminación, evitar la transpiración y pérdida de agua. La señal de sequía aumenta la cantidad endógena de algunas fitohormonas como son el ácido Abscisico y/o el Etileno, ambas determinantes de efectos que aceleran la senectud de las hojas y promueven su caída más temprana. -
Las plantas perciben el verano, o la cercanía del periodo estacional de sequía. La señal de inicio de la floración no estaría sólo dada por el fotoperiodo sino por la percepción de incremento de temperaturas y de agua disponible, que asegure completar la fase de desarrollo reproductivo. La fenología de las flores y su longevidad depende de la temperatura y disponibilidad de agua; con lo cual en años secos se muestra una reducción del crecimiento vegetativo (tallos más cortos) aunque la floración, que implica reproducción a través de semillas, puede verse menos afectada.
-
Existen otras estrategias que igualmente permiten la vida de plantas en ambientes de sequía extremos. Por ejemplo, el tamarugo (Prosopis tamarugo) que crece en la porción desértica de la "Pampa del Tamarugal". Se trata de una planta leguminosa que a primera vista no posee ninguna estrategia adaptativa y es del tipo de fijación de carbono C3. La estrategia de supervivencia del tamarugo es tener raíces pivotantes muy largas que dan cuenta del mayor volumen en cuanto al suministro de agua profunda, y otras más superficiales captan el agua generada por condensación durante la noche. El intenso crecimiento de la raíz pivotante es de origen genético. Se trata sin embargo de estrategias de alto riesgo, por lo extremo de las condiciones existentes que impiden la existencia de otras especies vegetales.
Otras estrategias inherentes a su adaptación desértica vinculadas a aspectos nutricionales y que también han logrado subsanar. La falta de agua superficial, especialmente de lluvias y baja humedad relativa, no es posible la degradación de biomasa aérea por microorganismos, limitando el reciclaje orgánico y acceso de nutrientes hacia las raíces, especialmente de nitrógeno; limita la disponibilidad de iones por parte de las raíces. La solución del problema es variada; la falta de nitrógeno en las capas profundas se suple por fijación del N2 del aire, gracias a la asociación de las raíces de estas leguminosas con Rhizobium, bacteria que vive en forma simbiótica y que aporta este elemento en forma de amina evitando las deficiencias nutricionales previstas. Tamarugos cuentan además con un sistema superficial de raíces el cual alterna flujos hídricos que permite disolver y captar nutrientes superficiales.
Bosque de Tamarugo.
-
Las plantas manejan mecanismos fisiológicos específicos de protección. Algunas plantas sometidas a estrés hídrico y/o alta radiación (exceso de fotones) pueden contar con mayor resistencia y/o protección mediante una adecuada neutralización de las formas reactivas del O2, referidas a "especies reactivas del oxígeno (ROS)". La reactividad del ozono (O3) con O2 produce las ROS en términos de H2O2, cada uno de los cuales pueden producir daño por oxidación en genes alterando la configuración aminoacídica de proteínas. El O3 es altamente reactivo y puede producir oxidación de ácidos grasos a peróxidos lipídicos alterando la fluidez de las membranas; afectar las membranas tilacoidales de los sistemas pigmentarios de la fotosíntesis, dañar la enzima Rubisco, descontrolar los mecanismos de apertura y cierre de los estomas, entre otros efectos. En plantas en que concurre este ciclo protector éstas se encontrarán más protegidas en ambientes donde exista gran polución, especialmente en zonas urbanas e industriales con incrementos notorios de niveles de ozono (O3) en la atmósfera.
Junto a una alta radiación, se ha visto que en condiciones ambientales de continuas altas temperaturas, se induce la síntesis de "Proteínas de Shock Térmico" (HSPs) en muchas especies. Estas proteínas no sólo conceden una protección inmediata a altas temperaturas sino simultáneamente apoyan otros tipos de estrés. En general, las plantas que deben someterse a muy bajas temperaturas y están ambientadas a inviernos largos y fríos cuentan con estrategias diversas; entre ellas una proporción mayor de ácidos grasos insaturados vs. saturados en membranas de mitocondrias y membranas celulares; y acumulación de solutos en invierno para impedir la cristalización del agua a nivel tisular.
TOLERANCIA A LA INUNDACION Y O CONJUNTAMENTE A AGUAS SALINAS
La concentración de NaCI en el agua de mar, y su ion Na, implica altamente toxicidad para las células radicales alterando la estructura y funcionamiento de las membranas plasmáticas, lo que afecta su permeabilidad y capacidad de regulación. Existen varios mecanismos alternos, complementarios. Primeramente, en el caso de manglares, las plantas no son de gran altura y presentan un crecimiento lento. La mayoría de sus raíces están sumergidas en el agua crecen en contra de la gravedad y emergen por sobre el agua como zancos. La emergencia de las raíces es a consecuencia de la necesidad de absorber O2 para usar este gas en la combustión aeróbica en mitocondrias para la producción de ATP. El ATP es requerido para regular el ingreso excesivo de sales; regulándose así calidad y cantidad de sales que ingresan. El mecanismo constituye trabajo de bombas de cationes para facilitar el movimiento de iones hacia fuera de la célula en contra del gradiente externo más concentrado del ambiente, en este caso el agua de mar.
En resumen, para que el agua fluya desde el suelo a la atmósfera, atravesando haces vasculares, debe existir un gradiente de "potencial de agua"; mayor en el suelo y en la zona radical y menor en la porción foliar, donde se evapora. Para mantener dicho gradiente, las sales son filtradas no ingresando a la corriente transpiratoria; en otros casos, parte de las sales son excretadas por los tallos u hojas (glándulas de sal) y así eliminadas del sistema.
Existen otros aspectos metabólicos necesarios para mantener el equilibrio de plantas adaptadas a ambientes inundados, como por ejemplo una enzimática especial en las raíces. Uno de los mecanismos consiste en la actividad de la PEP-carboxilasa, sólo presente o activa en las raíces de plantas tolerantes a la inundación, capta el CO2 del sistema respiratorio y lo transforma en ácido málico estable, molécula usada posteriormente en síntesis de aminoácidos.
TOLERANCIA A VARIOS GASES
Las plantas mantienen un intenso metabolismo gaseoso debiendo regular concentraciones de gases producidos en respuesta a procesos endógenos o de origen externo. Los gases son los elementos más difundibles en los tejidos, penetrando membranas con gran facilidad y velocidad. Cutina, corteza y peridermis son respuestas anatómicas adaptativas previstas para regular niveles de gases (y agua) en hojas, tallos y raíces respectivamente.
-
Gases vinculados a fotosíntesis. La toxicidad para las plantas puede ocurrir cuando existen cambios internos bruscos de niveles de ambos gases. A veces puede existir una deficiencia de CO2 que provocaría la muerte por "desnutrición" de carbohidratos. Algunas especies de geófitas (entre ellas Stylites, que vive enterrada en el suelo), no cuentan con estomas en sus hojas. Dado a su hábito sub-superficial, la naturaleza le ha dado la adaptación consistente en que el suministro de CO2 sea incorporado por la raíz y se conduzca de allí al follaje para su fijación fotosintética.
-
Gases involucrados en regulación del desarrollo. En el caso particular de muchas plantas que viven en ambientes de suelos de humedad normal y que temporalmente deben soportar inundaciones naturales, ya sea por desborde de ríos, precipitaciones intensas o regadíos no controlados, éstas reaccionan produciendo el gas etileno en sus hojas. Por causa de inundación y anaerobiosis (y otros factores de estrés), se sintetiza en las raíces el precursor de etileno ACC a partir del aminoácido metionina, el cual se moviliza por el xilema a las hojas transformándose ahí en gas "etileno". Su acumulación provoca la caída total de hojas de la planta. El etileno no sólo se produce por causa de inundación sino que es una hormona natural endógena reguladora de la maduración en muchas especies. Aplicaciones al follaje deben ser por tanto cuidadosas para evitar una posible caída total de frutas con pérdida absoluta de cosechas y considerables daños en la vegetación que se encuentra aledaña.
-
Gases de naturaleza ajena a las plantas y su tolerancia. Otra situación particular referida a "gases" son posibles daños generados por estos contaminantes derivados del proceso industrial en áreas industriales. Otro ejemplo, serían los pesticidas, ya que suponen para el vegetal un estrés químico importante y este se prepara para degradarlos gracias a las proteínas “antiestrés”, o echarlos fuera mediante la rotura de las vacuolas.
-
TOLERANCIA A LA RADIACIÓN
Cuando la intensidad de fotones que llega a la superficie de la hoja es muy alta y se produce un exceso de fotones no utilizables en la fotosíntesis. Debe recordarse además que en términos evolutivos, la captación de fotones está ajustada. Plantas de sombra y de sol en su hábitat tienen diferente número de cloroplastos en sus células lo que permite regular la radiación con eficiencia; cambios de su orientación facilitan además ajustes rápidos según la radiación imperante del momento. También las hojas enfrentan la radiación con ajustes de posición, los movimientos pueden corresponder a respuestas de irritabilidad diurnas reversibles o bien a una adaptación continua de posición.
A nivel celular, parte de los fotones en exceso se eliminan como calor sin mayor problema; sin embargo, a nivel de la membrana, el exceso de fotones provoca daño de estructura el cual debe ser reparado. Como se indicó antes, a pesar que existen sistemas de protección, existen daños debido a la producción de diferentes formas tóxicas de oxígeno. Existe adicionalmente otro tipo de protección a nivel de los sistemas pigmentarios. Acá las formas tóxicas del oxígeno y compuestos derivados pueden ser reducidas o eliminadas por efecto de carotenoides y enzimas, pero en caso extremo, la proteína D1 del foto-sistema pigmentario II debe aceptar dichos productos a costa de su propia destrucción. Aunque las proteínas D1 son fácilmente recuperables las etapas posteriores de daño ocurren igual dada la formación de moléculas oxidadas, con resultante de fotoinhibición. -
Revista “Creces”. Julio 2001. Universidad Diego Portales
-
“Biología”. Sanz Esteban M; Serrano Barrero S; Torralba Redondo B.
4. Aprovechamiento: aplicación médica y agrícola
-Aprovechamiento agrícola
El término biocontrol en agricultura puede definirse como: “el uso de organismos, o de sustancias producidas durante su crecimiento, para reducir o eliminar plagas que afecten a los cultivos”. Uno de los mecanismos más novedosos para realizar biocontrol, y con mayor proyección de futuro, consiste en la utilización de determinadas moléculas para estimular la resistencia de la planta frente a fitopatógenos .
La aplicación del ASM (bajas dosis de aspirina) a cultivos de interés agrícola está bien establecida, produciendo una activación de la SAR(resistencia sistémica inducida) de larga duración y protegiendo a los cultivos contra agentes patógenos con un espectro específico a cada cultivo. También se ha demostrado la operatividad del empleo mixto de ASM con bajas dosis de fungicidas o bactericidas convencionales. Este tipo de tratamientos, conlleva una activación de los mecanismos de defensa de la planta así como una acción directa de los fungicidas contra los patógenos, lo que redunda en una disminución de las dosis requeridas de fungicidas o bactericidas, y unas prácticas agrícolas más sostenibles.
Hasta el momento, hemos descrito como todas las plantas responden al ataque de patógenos mediante diversas estrategias coordinadas de resistencia mediadas por diferentes rutas de señalización bioquímica. La rapidez y la extensión de la respuesta a éstas señales intracelulares, en la interacción planta-patógeno, determina finalmente, el que la planta pueda superar satisfactoriamente la infección del patógeno. En este sentido, la idea de acelerar la respuesta de la planta mediante la aplicación de inductores de resistencia sistémica (bióticos o abióticos, respetuosos con el medio ambiente) es del todo atractiva y supondría, al mismo tiempo, una alternativa biológica, ambiental y comercialmente viable, a los métodos actuales de control de patógenos mediante el uso tradicional de pesticidas químicos.
La demanda social de elicitores de respuestas de defensa de las plantas y compatibles con el medio ambiente, es cada vez mayor. En este sentido, compuestos abióticos como el ASM y Probenazol(fungicida sistémico de acción indirecta), o bióticos, considerados como compuestos compatibles con el medioambiente y se comercializan como compuestos que mejoran la resistencia/tolerancia de las plantas a enfermedades producidas por patógenos. Recientemente, se ha desarrollado un producto compuesto fundamentalmente por peróxido de hidrógeno OxycomTM , que al aplicarse sobre los cultivos en experimentos de campo y en el laboratorio, ha demostrado que induce la expresión de genes relacionados con las respuestas sistémicas de defensa a un buen nivel.
Una estrategia agrícola sostenible o ecológica, basada en la utilización de compuestos elicitores de mecanismos de defensa y que tenga como objetivo final la consecución de una inmunidad fisiológica en la planta, necesita para su aplicación el disponer en el mercado de una amplia gama de dichos productos. Por ello, una de las prioridades actuales de las empresas relacionadas con la fabricación de compuestos fitosanitarios, es la obtención de moléculas elicitoras de las respuestas sistémicas de defensa de las plantas que sean susceptibles de ser utilizadas en una agricultura más respetuosa con el medio ambiente. Entre los factores positivos relacionados con el empleo de este tipo de compuestos, se podrían destacar los siguientes :
1) Evitan los problemas relacionados con la aparición de resistencias en los patógenos a los pesticidas clásicos.
2) Pueden sustituir a los numerosos pesticidas que actualmente son de retirada obligatoria, al no ser compatibles medioambientalmente.
3) Reducen los riesgos asociados a la salud que pueden presentar los pesticidas químicos.
4) Pueden ser efectivos contra enfermedades producidas por virus y patógenos de suelo, frente a los cuales los tratamientos químicos no son eficaces.
5) No hay necesidad de introducir genes foráneos en las plantas, y por tanto se evita el problema-dilema de los transgénicos.
6) Presenta un efecto de larga duración y de amplio espectro.
7) Se activan defensas naturales de la planta mediante la inducción de resistencia, evitando se generen resistencias indeseadas en los patógenos.
Sin embargo, este tipo de estrategia también tiene sus inconvenientes. Entre los inconvenientes a considerar se podrían citar :
1) Al contrario de los pesticidas clásicos que actúan destruyendo o inhibiendo el crecimiento de los patógenos, la resistencia sistémica inducida depende de la expresión de genes en la planta, su efectividad estando sujeta a influencias fisiológicas y medioambientales.
2) Las compañías sólo comercializarían elicitores patentables.
3) La aprehensión de los productores y consumidores por las nuevas tecnologías.
Uno de los principales problemas con los que nos enfrentamos a la hora de producir o generar nuevos compuestos elicitores, es el de contar con técnicas que permitan detectar su efecto en la planta de una forma rápida y fiable. Los ensayos que se realizan con este fin, en la actualidad, se basan en pruebas de campo (alto consumo de tiempo) y/o en la detección de productos resultantes de la inducción de los mecanismos de defensa de la planta (alto consumo de tiempo y pueden ser inespecíficos). En ambos ensayos es muy complicado determinar el tipo e intensidad de respuesta de la planta. Por lo tanto, para poder realizar una búsqueda sistemática de nuevos compuestos elicitores, es imprescindible contar con técnicas que permitan de forma fiable y rápida, constatar y cuantificar la respuesta de la planta frente a los mismos.
La novedad de utilizar fermentaciones microbianas a la hora de generar nuevos elicitores se basa, en primer lugar, en que los microorganismos a emplear se caracterizan por una gran capacidad metabólica y de secreción de enzimas extracelulares, lo cual nos asegura que durante su crecimiento e hidrólisis de los sustratos de fermentación, se van a liberar y/o generar una amplia gama de sustancias a los medios de cultivo. Por otro lado, los subproductos y residuos que se emplean como sustratos de fermentación, han sido escogidos por ser fuentes de macromoléculas, que una vez hidrolizadas, podrían dar lugar a compuestos elicitores, como por ejemplo: quitina, glucanos, xilanos, pectinas, proteínas, lignina, etc. A lo anteriormente mencionado, habría que añadir que los sustratos son de fácil disponibilidad y bajo costo.
La combinación de ambos procesos, producción y escrutinio o “screening” de compuestos elicitores mediante la tecnología molecular antes descrita, pese a su reciente puesta a punto, nos está permitiendo alcanzar resultados muy prometedores, obteniéndose productos de superior actividad elicitora que los compuestos actualmente comercializados.
-Aplicaciones médicas.
En los procesos biológicos las proteasas ocupan un lugar altamente relevante, ya que son ellas las encargadas de procesar las pro enzimas (zimógenos) , digerir los receptores para evitar su reconocimiento, controlar la duración de péptidos bioactivos, controlar la existencia de proteínas defectuosas, digerir las proteínas de los alimentos, entre otras muchas funciones. En las células patológicas como es el caso de las cancerígenas las proteasas juegan un papel determinante, tanto en el crecimiento de tumores, como en su vascularización (angiogénesis) y al mismo tiempo son un requisito indispensable para poder realizar el fenómeno de metástasis. De hecho, son ellas las que permiten liberar ala ulula de su entorno, para poder ingresar al torrente circulatorio e iniciar el cáncer. Este fenómeno esta regido por un fino balance entre las proteasas existentes en la célula, las proteasas que excreta al medio y sus respectivos inhibidores naturales que las mismas células producen. Cuando este balance se altera (estado patológico) y las proteasas actúan sobre las proteínas de la matriz extracelular, se inicia el proceso de liberación de células malignas, iniciándose así el cáncer. Es aquí donde los inhibidores de proteasas extraídos de las plantas , pueden jugar un papel terapéutico, al inhibir la actividad de esas proteasas, impidiendo así su liberación y por tanto la metástasis.
Probando los distintos inhibidores que hemos aislado de las distintas plantas, encontramos que tanto el inhibidor de chan como el del fríjol tépari disminuían el avance de las células malignas, principalmente el inhibidor del fríjol tépari, el cual mostró un importante efecto sobre algunos cultivos de líneas celulares cancerígenas, impidiendo la liberación de las células.
Este efecto fue selectivo y dependiente de la concentración, detectándose a dosis muy bajas de este compuesto. Actualmente se está determinando sobre que espectro de líneas celulares actúa, así como sobre cuáles son las proteasas blanco sobre las que ejerce su efecto, a fin de entender su mecanismo molecular reacción en ese importante proceso patológico.
Finalmente se ha trabajado con otras proteínas también relacionados con los mecanismos de defensa de las plantas, como son las lectinas de las cuales hemos detectado que también tienen una actividad sobre células cancerígenas, en este caso disminuyendo la proliferación celular. Esta molécula también participa en la defensa de la planta, a través de los mecanismos de oxidación, propiciando la formación de especies reactivas al oxígeno.
5. Comportamiento de plantas transgénicas de tabaco contra patógenos e insectos
Las plantas para sobrevivir han tenido que desarrollar la capacidad de adaptarse a un medio hostil, mediante la expresión de una gran diversidad de respuestas, que les permiten hacer frente a algún agresor en situaciones de agobio ambiental. Estos sistemas de defensa incluyen proteínas, moléculas de señal y metabolitos secundarios involucrados no sólo en defensa, sino en repelencia, tolerancia y demás.
Cuando ocurre un daño mecánico o herida en la planta, se desencadenan una serie de eventos bioquímicos y moleculares idénticos a los que ocurren como consecuencia del ataque de insectos.
La respuesta de un gen a una herida, por tanto es una indicación de la respuesta de defensa al ataque por insectos. Los peroxisomas están relacionados con la producción y degradación del peróxido de hidrógeno, pero también son los organelos encargados de degradar ácidos grasos y dependiendo de su localización pueden ser separados en glioxisomas, peroxisomas de hojas y no especializados. Los glioxisomas juegan un papel importante en la conversión de lípidos a carbohidratos en la beta- oxidación, misma ruta que es inducida por una serie de factores de transcripción conocidos como receptores activados de proliferadores peroxisomales (PPAR), y que inducen la expresión de genes implicados en la beta -oxidación.
Se trabajó con líneas transgénicas de tabaco, T2 y T5, en las que se insertó el gen que codifica para los genes receptores activadores de la proliferación de peroxisomas (PPAR), para determinar posibles similaridades en los mecanismos de regulación en la proliferación peroxisomal de plantas y animales. Dichas plantas transgénicas tuvieron cambios morfológicos en las hojas y hubo una gran variación en el contenido de ácidos grasos de las transgénicas con respecto al tipo silvestre, donde en general los ácidos grasos saturados fueron más abundantes en los transgénicos y los insaturados más deficientes, y muchos ácidos grasos no fueron detectados. La línea T2 fue la que presentó los mayores cambios en el tamaño de la hoja, siendo más grande y mucho más corrugada con respecto al tipo silvestre que las otras líneas, además de presentar floración mucho antes que los otros fenotipos. Pero entonces, para qué determinar similaridades entre los mecanismos de regulación en la proliferación peroxisomal entre plantas y animales, pues porque hay publicaciones de artículos que demuestran que existe una interrelación entre las diferentes rutas de defensa de las plantas y los ácidos grasos, ya que estos son precursores de importantes moléculas de señal. Y en este caso resalta el ácido linolénico (18:3) como precursor de las mismas, divido a la gran variación que muestra en las líneas transgénicas.
Entonces, bajo este contexto, se planteó el propósito de ver cómo responden éstas líneas transgénicas, con el metabolismo de los ácidos grasos alterados, a la agresión contra bacterias patógenas e insectos. Para ver si existía una respuesta diferencial al ataque contra bacterias patógenas se diseñó un experimento con distintos organismos que se sabe atacan al tabaco, y lo que se hizo fue infiltrar en la 4ª hoja de las plantas de tabaco una solución de un buffer de NaPO4 y bacterias de aproximadamente 0.3 unidades de absorbancia para cada bacteria y un tercer grupo de plantas como control con puro buffer. Para el ensayo de resistencia contra insectos, se diseño un experimento con mosquita blanca, en el que se aislaban en jaulas una planta de cada línea y el control. A cada planta previamente se le pusieron 25 mosquitas adultas con una “clip cage” para que ovipositaran en una sola hoja y se dejaron 36 horas a 27 º C, luego se retiraron las mosquitas adultas y las plantas se mantuvieron aisladas por 17 días, mientras el 70 % eclosionaban con el propósito de ver sí hay alguna diferencia en el número de mosquitas eclosionadas o sin eclosionar en cada hoja infestada.
Resultados del ensayo con bacterias.
Las plantas de tabaco inoculadas con P. s. helianthis se necrosaron completamente 3 días después de la infiltración y no dio tiempo de medir ninguna reacción diferencial entre las líneas. Por otro lado, las bacterias infiltradas con P. s. Tagetis presentaron una reacción hipersensible que es una reacción de respuesta rápida en la que la planta mata todas las células de alrededor de la lesión o infección evitando que el patógeno se disemine.
En las plantas infectadas con P. s. Tagetis se midió el área de la lesión en las líneas transgénicas y el control y se comparó contra las mismas líneas inoculadas con el buffer y se encontró que el área de lesión entre el control y T5 infectados con Tagetis no tuvieron diferencia entre sí, pero sí la hubo contra T2. Y entre las tres líneas inoculadas con buffer no hubo diferencias significativas, sin embargo sí presentaron un área de lesión mucho menor con respecto a las que fueron infectadas.
Resultados del ensayo con mosquita blanca.
En este ensayo, se contaron el número de mosquitas eclosionadas contra el número de mosquitas no eclosionadas, y se encontró que entre las plantas control y T2 no hubo diferencia significativa, sin embargo sí la hubo con respecto al número de mosquitas eclosionadas en T5 que fue mucho menor.
Una vez obtenidos los resultados del ensayo de las plantas inoculadas con P. s. tagetis y con el buffer de NaPO4, se pulverizó el tejido infectado en nitrógeno líquido y se extrajo el RNA para hacer un northern, técnica de electroforesis que consiste en la separación de las diferentes secuencias de ARN por su peso molecular o tamaño. Se corrió un gel de agarosa desnaturalizante con el RNA para separarlo y se transfirió a una membrana de nitrocelulosa a la que se agrega una sonda radiactiva. La sonda elegida fue la ACCO, ya que es un metabolito secundario en la ruta de defensa de las plantas y su presencia indicaría que las líneas transgénicas presentan una reacción ante al ataque por patógenos.
0.00 20.00 40.00 60.00 80.00 100.00
Tc
T2
T5
% Eclosionadas X % No eclosionadas X
0.0
0.2
0.4
0.6
0.8
1.0
Tagetis Buffer
ÁREAS
Tc
T2
T5
Esta placa muestra que hay expresión del gen de la ACCO en todas las plantas, pero es mucho menor en la línea T2. A partir de los resultados obtenidos en los anteriores ensayos, pude concluir que sí hay una respuesta diferencial en las diferentes líneas transgénicas de Tabaco, debido a la alteración de su metabolismo de los ácidos grasos.
6. Investigaciones:
6.1. Coevolución plantas-animales
Las mariposasHeliconius se alimentan casi en forma exclusiva de polen y néctar de las flores de Passiflora (género de plantas a las que pertenece por ejemplo nuestra enredadera Mburucuyá o Pasionaria).
Estas plantas producen sustancias de las que deriva el cianuro para protegerse de los insectos herbívoros. Pero las Heliconius lo pueden procesar. Sus orugas lo ingieren al comer las hojas y sintetizan su propio veneno, mientras que las mariposas adultas lo obtendrían a partir del polen. El predador desprevenido con capacidad de aprendizaje (como las aves) que capture una de estas mariposas sabrá que no debe intentarlo nuevamente. Recordará los colores de las Heliconius que actúan como una gran señal luminosa advirtiendo: "no intenten comerme, soy tóxica". Otros géneros de mariposa simplemente "copian" los colores, favoreciéndose de esta característica.
En el sector Cataratas del Parque Nacional Iguazú el Ambay (Cecropia pachistachya) es uno de los árboles más frecuentes. Tienen el tronco recto con compartimentos huecos separados por tabiques donde viven hormigas del género Azteca. Las agresivas hormigas protegen su territorio, en este caso el árbol, contra diversos herbívoros como las hormigas podadoras. A cambio obtienen refugio y alimento de estructuras especiales llamadas cuerpo de Mullerian, que son paquetes de hidratos de carbono situados en la axila de las hojas.
Muchas especies utilizan las hormigas para defensa atrayéndolas con estructuras glandulares llamadas nectarios extraflorales. Se ubican en cualquier parte de la planta que no sean las flores, de allí su nombre. La producción de néctar, líquido azucarado, atrae a las hormigas, las que protegen a la planta (su alimento) de otros predadores. (Foto: nectarios extraflorales en hoja de Turnera ulmifolia)
Muchas especies aprovechan la visita de las hormigas para la dispersión de las semillas. Para ello las semillas presentan estructuras atrayentes como carúnculas y arilos. La carúncula de la semilla del Ricino es una excrescencia del tegumento externo de la semilla rico en aceites (elaiosomas). El arilo está presente en numerosos géneros (Lychnis), tienen abundantes aceites y almidones y se forma a partir de diferentes partes de la semilla.
6.2. Científicos descubren un gen que inmuniza a las plantas contra los hongos
El grupo científico que dirige el profesor del departamento de Biotecnología de la Universidad Politécnica de Madrid (UPM), Antonio Molina, junto con investigadores alemanes y estadounidenses, ha identificado un componente básico de inmunidad de las plantas a los hongos patógenos, hasta ahora desconocido.
El descubrimiento de este gen de resistencia, al que los investigadores han denominado PEN2 (penetración), supone "un gran paso dentro del conocimiento del mecanismo de defensa de las plantas", explicó Molina.
Este gen resistente fue identificado en la planta Arabidopsis thaliana, una especie modelo en la que el grupo de investigación del Departamento de Biotecnología de la UPM ha centrado sus investigaciones, publicadas en el último número de la revista Science. Según explicó, las plantas se exponen a gran cantidad de patógenos de los que sólo un reducido número es capaz de infectar una especie vegetal, y ello significa que la planta presenta una resistencia de "no huésped" cuando el patógeno es incapaz de infectarla.
Hasta el momento se creía que esta resistencia de una planta a los patógenos dependía de mecanismos pasivos como la estructura de la pared celular o la existencia de sustancias antimicrobianas en su superficie, pero se ha demostrado que contribuye una respuesta inmune activa.
"Hasta este momento se pensaba que era el patógeno el que no era eficiente cuando no conseguía infectar a una planta", explicó Molina.
Ahora se sabe que "las plantas tienen un mecanismo de defensa basal", además de "varias etapas de defensa con barreras consecutivas", que el patógeno debe superar y que determinan si la planta es susceptible o resistente.
Según los investigadores, la pérdida de función de PEN2 hace que las plantas sean más susceptibles a enfermedades causantes de graves en cosechas, como el mildiu de la patata, o determinados hongos necrotrofos, y de ello se deduce que es un componente básico del sistema de inmunidad innata de las plantas.
Entre las aplicaciones de este descubrimiento destaca la posibilidad de la utilización de estos componentes en el desarrollo de fungicidas naturales o "verdes", con un amplio espectro de acción frente a diferentes enfermedades.
GLOSARIO:
Plantas C3: son las plantas cuyo sistema fotosintético de fijación de CO2 se produce exclusivamente con la enzima Rubisco que toma el CO2 y lo liga a una pentosa, siendo el primer producto de fotosíntesis una triosa, es decir un azúcar de 3 carbonos (3-fosfoglicerato); en consecuencia C3.
Plantas CAM o MAC: son aquellas que poseen además de la Rubisco la enzima PEP-carboxilasa. La PEP-carboxilasa capta en la noche, a estoma abierto, el CO2 y lo fija a malato. El malato se acumula en altas concentraciones en la vacuola como ácido málico bajando el pH (acidificando el jugo vacuolar, de allí metabolismo ácido.) Durante el día, a estoma cerrado, el acido málico se decarboxila (se hace menos ácido) y el CO2 antes fijado se une ahora por la Rubisco a una pentosa en el estroma del cloroplasto formando el glicerato característico resultante de la reacción.
Rubisco: enzima 1,5-difosfatocarboxilasa/oxidasa. Corresponde más o menos al 50% de las proteinas presentes de una hoja. La enzima capta el CO2 disponible en el Ciclo de Calvin formando 3-fosfoglicerato. Sin embargo, es capaz de aceptar también O2 y lo realiza en un ciclo llamado "fotorespiración" por el cual la eficiencia de fijación del CO2 en las plantas C3 puede ser relativamente baja a alta irradiación.
Especies reactivas de oxígeno (ROS): se refiere a los distintos cambios electrónicos y de carga que puede tomar el oxígeno; formas todas reactivas y que dañan la estabilidad celular. Ocurre cuando hay exceso de electrones que pueden ser donados al O2 a nivel de la cadena de transporte de electrones en mitocondrias y cloroplastos.
Proteína D1 del Sistema Pigmentario II: proteína de 32 kDa perteneciente a dicho sistema pigmentario envuelta en el transporte inicial de electrones derivado de la fotolisis del agua y en la producción de O2 como consecuencia de lo anterior.
Nematodos: fílum de gusanos unisexuales, de cuerpo cilíndrico y delgado, sin segmentar, y cubierto por una cutícula; existen formas libres y parásitas.
Micoplasma: microorganismo unicelular de acción patógena, de pared celular poco diferenciada y con gran variedad de formas.
Fungicida: agente que destruye los hongos.
Patogénesis: origen y desarrollo de las enfermedades.
Lisozima: es una enzima muy ampliamente distribuida entre los seres vivos, desde virus a eucariota.
Tubérculo: es un tallo subterráneo modificado y engrosado donde se acumulan los nutrientes de reserva para la planta.
Cisteìna: es uno de los veintidós aminoácidos que utilizan las células para sintetizar proteínas. Está codificada en el ARN mensajero como UGU o UGC.
Quitosana: derivados de quitina.
PEN: enzima localizada en la membrana de los peroxisomas.
Proteínas PR: proteínas implicadas en la patogénesis, que limitan la acción del patógeno de la planta.
PGIP: es una proteína secretada y localizada en la matriz extracelular.
BIBLIOGRAFÍA:
Portal web: www.creces.cl
Ed. Oxford. Proyecto EXEDRA.
21
Descargar
| Enviado por: | Winds |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.