Biología, Botánica, Genética y Zoología
Fisiología del sistema auditivo
Fisiología del
Sistema Auditivo
En este capítulo se examina la estructura y funcionamiento del oído, con el fin de lograr una mejor comprensión de los fenómenos y modelos psicoacústicos. Se estudia la anatomía y la fisiología del aparato auditivo, haciendo énfasis en aquellas partes y estructuras del mismo más importantes para el desarrollo de modelos perceptuales.
III.1.- El sentido de la audición y el sistema auditivo
La generación de sensaciones auditivas en el ser humano es un proceso extraordinariamente complejo, el cual se desarrolla en tres etapas básicas:
-
Captación y procesamiento mecánico de las ondas sonoras.
-
Conversión de la señal acústica (mecánica) en impulsos nerviosos, y transmisión de dichos impulsos hasta los centros sensoriales del cerebro.
-
Procesamiento neural de la información codificada en forma de impulsos nerviosos.
La captación, procesamiento y transducción de los estímulos sonoros se llevan a cabo en el oído propiamente dicho, mientras que la etapa de procesamiento neural, en la cual se producen las diversas sensaciones auditivas, se encuentra ubicada en el cerebro. Así pues, se pueden distinguir dos regiones o partes del sistema auditivo: la región periférica, en la cual los estímulos sonoros conservan su carácter original de ondas mecánicas hasta el momento de su conversión en señales electroquímicas, y la región central, en la cual se transforman dichas señales en sensaciones.
En la región central también intervienen procesos cognitivos, mediante los cuales se asigna un contexto y un significado a los sonidos [1]; es decir, permiten reconocer una palabra o determinar que un sonido dado corresponde a un violín o a un piano.
El presente trabajo se limita a estudiar y utilizar solamente los aspectos perceptuales del sistema auditivo; esto es, aquellos que son independientes del contexto y del significado y que, en buena parte, se localizan en la región periférica.
III.1.1 Región periférica del sistema auditivo
El oído o región periférica se divide usualmente en tres zonas, llamadas oído externo, oído medio y oído interno, de acuerdo a su ubicación en el cráneo, como puede verse en la Fig.III.1.
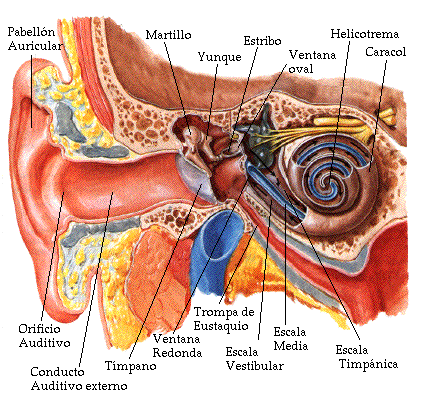
Fig.III.1. Anatomía del oído humano.
Los estímulos sonoros se propagan a través de estas zonas, sufriendo diversas transformaciones hasta su conversión final en impulsos nerviosos. Tanto el procesamiento mecánico de las ondas sonoras como la conversión de éstas en señales electroquímicas son procesos no lineales [2] [3], lo cual dificulta la caracterización y modelado de los fenómenos perceptuales.
En las siguientes secciones de este capítulo se estudia la anatomía y funcionamiento de estas tres zonas del oído, así como la propagación y procesamiento del sonido a través de las mismas.
III.2 Oído externo
III.2.1 Anatomía y funcionamiento
El oído externo (Fig. III.1) está formado por el pabellón auricular u oreja, el cual dirige las ondas sonoras hacia el conducto auditivo externo a través del orificio auditivo. El otro extremo del conducto auditivo se encuentra cubierto por la membrana timpánica o tímpano, la cual constituye la entrada al oído medio. La función del oído externo es la de recolectar las ondas sonoras y encauzarlas hacia el oído medio. Asimismo, el conducto auditivo tiene dos propósitos adicionales: proteger las delicadas estructuras del oído medio contra daños y minimizar la distancia del oído interno al cerebro, reduciendo el tiempo de propagación de los impulsos nerviosos [3].
III.2.2 Respuesta en frecuencia y localización de las fuentes de sonido
El conducto auditivo es un "tubo" de unos 2 cm de longitud, el cual influye en la respuesta en frecuencia del sistema auditivo. Dada la velocidad de propagación del sonido en el aire (aprox. 334 m/s), dicha longitud corresponde a 1/4 de la longitud de onda de una señal sonora de unos 4 kHz. Este es uno de los motivos por los cuales el aparato auditivo presenta una mayor sensibilidad a las frecuencias cercanas a los 4 kHz, como se verá en el siguiente capítulo.
Adicionalmente, el pabellón auricular, junto con la cabeza y los hombros, contribuye a modificar el espectro de la señal sonora. Las señales sonoras que entran al conducto auditivo externo sufren efectos de difracción debidos a la forma del pabellón auricular y la cabeza, y estos efectos varían según la dirección de incidencia y el contenido espectral de la señal; así, se altera el espectro sonoro debido a la difracción [4]. Estas alteraciones, en forma de "picos" y "valles" en el espectro, son usadas por el sistema auditivo para determinar la procedencia del sonido en el llamado "plano medio" (plano imaginario perpendicular a la recta que une ambos tímpanos) [1] [5].
III.3 Oído medio
III.3.1 Anatomía
El oído medio (Fig. III.2) está constituido por una cavidad llena de aire, dentro de la cual se encuentran tres huesecillos, denominados martillo, yunque y estribo, unidos entre sí en forma articulada. Uno de los extremos del martillo se encuentra adherido al tímpano, mientras que la base del estribo está unida mediante un anillo flexible a las paredes de la ventana oval, orificio que constituye la vía de entrada del sonido al oído interno.
Finalmente, la cavidad del oído medio se comunica con el exterior del cuerpo a través de la trompa de Eustaquio, la cual es un conducto que llega hasta las vías respiratorias y que permite igualar la presión del aire a ambos lados del tímpano.
III.3.2 Propagación del sonido y acople de impedancias
Los sonidos, formados por oscilaciones de las moléculas del aire, son conducidos a través del conducto auditivo hasta el tímpano. Los cambios de presión en la pared externa de la membrana timpánica, asociados a la señal sonora, hacen que dicha membrana vibre siguiendo las oscilaciones de dicha señal.
Las vibraciones del tímpano se transmiten a lo largo de la cadena de huesecillos, la cual opera como un sistema de palancas [6] [3], de forma tal que la base del estribo vibra en la ventana oval (ver la Fig. III.2). Este huesecillo se encuentra en contacto con uno de los fluidos contenidos en el oído interno; por lo tanto, el tímpano y la cadena de huesecillos actúan como un mecanismo para transformar las vibraciones del aire en vibraciones del fluido.
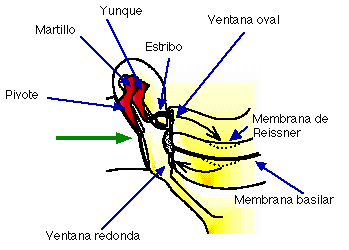
Fig. III.2. Propagación del sonido a través del oído medio e interno.
Ahora bien, para lograr que la transferencia de potencia del aire al fluido sea máxima, debe efectuarse un acoplamiento entre la impedancia mecánica característica del aire y la del fluido, puesto que esta última es mucho mayor que la primera.
Un equivalente mecánico de un transformador (el acoplador de impedancias eléctricas) es, precisamente, una palanca [3]; por ende, la cadena de huesecillos actúa como acoplador de impedancias. Además, la relación entre las superficies del tímpano y de la base del estribo (en la ventana oval) introduce un efecto de acoplamiento adicional, lográndose una transformación de impedancias del orden de 1:20 [4], con lo cual se minimizan las pérdidas por reflexión.
El máximo acoplamiento se obtiene en el rango de frecuencias medias, en torno a 1 kHz [3]. En la Fig. III.3 se representa en forma esquemática la transmisión del sonido del oído externo al interno, a través del oído medio.
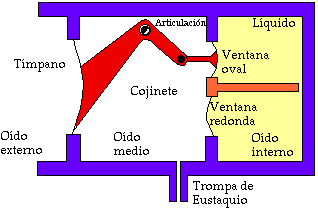
Fig. III.3. Esquema de la propagación del sonido a través del oído medio.
III.3.3 Reflejo timpánico o acústico
Cuando se aplican sonidos de gran intensidad (> 90 dB SPL) al tímpano, los músculos tensores del tímpano y el estribo se contraen de forma automática, modificando la característica de transferencia del oído medio y disminuyendo la cantidad de energía entregada al oído interno.
Este "control de ganancia" se denomina reflejo timpánico o auditivo, y tiene como propósito proteger a las células receptoras del oído interno frente a sobrecargas que puedan llegar a destruirlas. Este reflejo no es instantáneo, sino que tarda de 40 a 160 ms en producirse [6].
El reflejo timpánico debe ser tomado en cuenta en cualquier modelo matemático del procesamiento del sonido en el aparato auditivo, siempre que se trabaje con sonidos de gran intensidad [7], puesto que es un mecanismo no lineal que introduce un término cuadrático en la relación entrada-salida del oído medio [4].
III.3.4 Respuesta en frecuencia combinada del oído externo y el oído medio
El conjunto formado por el oído externo y el oído medio forman un sistema cuya respuesta en frecuencia es de tipo pasabajos [1], como se muestra en la Fig. III.4. En el intervalo cercano a los 4 kHz se observa un pequeño efecto de ganancia, debido a las características del conducto auditivo.
Esta respuesta sólo es válida cuando el sistema se comporta de modo lineal; es decir, cuando la intensidad del sonido no es muy elevada, para evitar que actúe el reflejo timpánico.
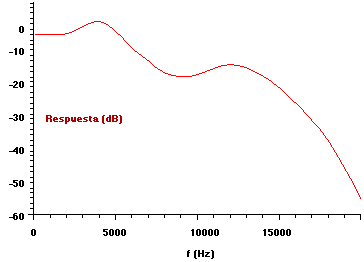
Fig. III.4. Respuesta en frecuencia combinada del oído externo y el oído medio
III.4 Oído interno
El oído interno representa el final de la cadena de procesamiento mecánico del sonido, y en él se llevan a cabo tres funciones primordiales: filtraje de la señal sonora, transducción y generación probabilística de impulsos nerviosos [8].
III.4.1 Anatomía
En el oído interno se encuentra la cóclea o caracol, la cual es un conducto rígido en forma de espiral (ver la Fig. III.1) de unos 35 mm de longitud, lleno con dos fluidos de distinta composición.
El interior del conducto está dividido en sentido longitudinal por la membrana basilar y la membrana de Reissner, las cuales forman tres compartimientos o escalas (Fig. III.5). La escala vestibular y la escala timpánica contienen un mismo fluido (perilinfa), puesto que se interconectan por una pequeña abertura situada en el vértice del caracol, llamada helicotrema. Por el contrario, la escala media se encuentra aislada de las otras dos escalas, y contiene un líquido de distinta composición a la perilinfa (endolinfa).
La base del estribo, a través de la ventana oval, está en contacto con el fluido de la escala vestibular, mientras que la escala timpánica desemboca en la cavidad del oído medio a través de otra abertura (ventana redonda) sellada por una membrana flexible (membrana timpánica secundaria).
Sobre la membrana basilar y en el interior de la escala media se encuentra el órgano de Corti (Fig. III.6), el cual se extiende desde el vértice hasta la base de la cóclea y contiene las células ciliares que actúan como transductores de señales sonoras a impulsos nerviosos. Sobre las células ciliares se ubica la membrana tectorial, dentro de la cual se alojan las prolongaciones o cilios de las células ciliares externas.
Dependiendo de su ubicación en el órgano de Corti, se pueden distinguir dos tipos de células ciliares: internas y externas. Existen alrededor de 3500 células ciliares internas y unas 20000 células externas [6]. Ambos tipos de células presentan conexiones o sinapsis con las fibras nerviosas aferentes (que transportan impulsos hacia el cerebro) y eferentes (que transportan impulsos provenientes del cerebro), las cuales conforman el nervio auditivo. Sin embargo, la distribución de las fibras es muy desigual: más del 90% de las fibras aferentes inervan a las células ciliares internas, mientras que la mayoría de las 500 fibras eferentes inervan a las células ciliares externas [6] [3]. El propósito de ambos tipos de células y de la distribución de las conexiones nerviosas se estudia más adelante, en la sección III.6, "Mecanismo de transducción".
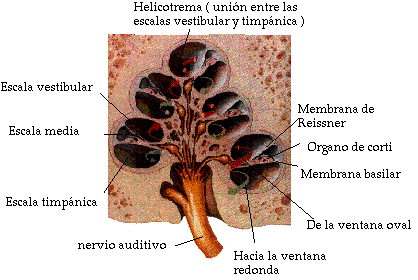
Fig. III. 5. Corte transversal de la cóclea o caracol.
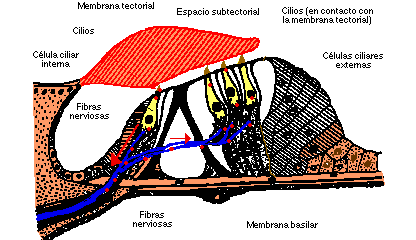
Fig. III.6. Órgano de Corti.
III.4.2 Propagación del sonido en la cóclea
Las oscilaciones del estribo (ver la Fig. III.2) provocan oscilaciones en el fluido de la escala vestibular (perilinfa). La membrana de Reissner, la cual separa los fluidos de la escala vestibular y la escala media, es sumamente delgada y, en consecuencia, los líquidos en ambas escalas pueden tratarse como uno solo desde el punto de vista de la dinámica de los fluidos [3]. Así, las oscilaciones en la perilinfa de la escala vestibular se transmiten a la endolinfa y de ésta a la membrana basilar (Fig. III.7); la membrana basilar, a su vez, provoca oscilaciones en el fluido de la escala timpánica.
Puesto que tanto los fluidos como las paredes de la cóclea son incompresibles, es preciso compensar el desplazamiento de los fluidos; esto se lleva a cabo en la membrana de la ventana redonda, la cual permite "cerrar el circuito hidráulico" [9].
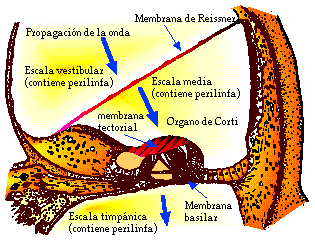
Fig. III.7. Corte transversal de un conducto de la cóclea.
La propagación de las oscilaciones del fluido en la escala vestibular a la timpánica no sólo se lleva a cabo a través de la membrana basilar; para sonidos de muy baja frecuencia, las vibraciones se transmiten a través de la abertura situada en el vértice de la cóclea (helicotrema).
En conclusión, el sonido propagado a través del oído externo y medio llega hasta la cóclea, donde las oscilaciones en los fluidos hacen vibrar a la membrana basilar y a todas las estructuras que ésta soporta.
III.5.La cóclea como analizador en frecuencia
La membrana basilar es una estructura cuyo espesor y rigidez no es constante: cerca de la ventana oval, la membrana es gruesa y rígida, pero a medida que se acerca hacia el vértice de la cóclea se vuelve más delgada y flexible.
La rigidez decae casi exponencialmente con la distancia a la ventana oval; esta variación de la rigidez en función de la posición afecta la velocidad de propagación de las ondas sonoras a lo largo de ella [5] [9], y es responsable en gran medida de un fenómeno muy importante: la selectividad en frecuencia del oído interno.
III.5.1.Ondas viajeras y transformación de frecuencia a posición
Las ondas de presión generadas en la perilinfa a través de la ventana oval tienden a desplazarse a lo largo de la escala vestibular. Debido a que el fluido es incompresible la membrana basilar se deforma, y la ubicación y amplitud de dicha deformación varía en el tiempo a medida que la onda de presión avanza a lo largo de la cóclea.
Para comprender el modo de propagación de las ondas de presión, supóngase que se excita el sistema auditivo con una señal sinusoidal de una frecuencia dada:
La membrana basilar vibrará sinusoidalmente, pero la amplitud de la vibración irá en aumento a medida que se aleja de la ventana oval (debido a la variación en la velocidad de propagación), hasta llegar a un punto en el cual la deformación de la membrana basilar sea máxima; en ese punto de "resonancia", la membrana basilar es acústicamente "transparente" (es decir, se comporta como si tuviera un orificio) [5], de modo que la amplitud de la vibración y, por ende, la transmisión de la energía de la onda al fluido de la escala timpánica es máxima en dicho punto.
A partir de esa región, la onda no puede propagarse eficientemente [9], de modo que la amplitud de la vibración se atenúa muy rápidamente a medida que se acerca al helicotrema. En la Fig. III.8 se observa la onda en la membrana basilar en un instante de tiempo.
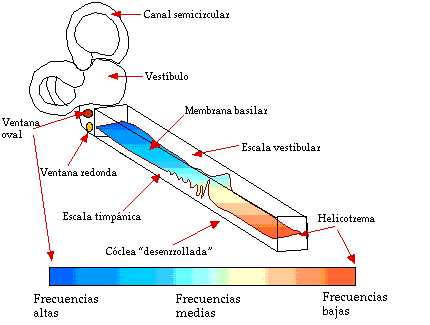
Fig. III.8. Onda viajera en la membrana basilar.
En este modo de propagación, las ondas de presión son ondas viajeras , en las cuales (a diferencia de las ondas estacionarias) no existen nodos [3]. En la Fig. III.9 se observa la amplitud de oscilación de la membrana basilar en dos instantes de tiempo, junto con la envolvente de la onda viajera, en función de la distancia al estribo.
La ubicación del máximo de la envolvente de la onda viajera depende de la frecuencia de la señal sonora, como puede observarse en la Fig. III.10: mientras menor es la frecuencia del tono, mayor es la distancia que viaja la onda a lo largo de la membrana antes de ser atenuada, y viceversa. De esta forma, la membrana basilar dispersa las distintas componentes de una señal de espectro complejo en posiciones bien definidas respecto a la ventana oval [3].
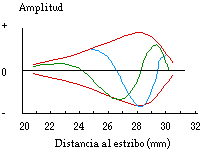
Fig. III.9. Ondas viajeras para un tono de 200 Hz.
Fig. III.10. Transformación de frecuencia a posición en la membrana basilar.
III.5.2.Selectividad en frecuencia de la membrana basilar
Como se ha visto, las altas frecuencias contenidas en un estímulo sonoro se atenúan a medida que la onda se desplaza hacia el helicotrema. Así, se puede considerar a la membrana basilar como un filtro pasabajos de parámetros distribuidos [9]. Por otro lado, si se midiese la respuesta en frecuencia en un punto dado de dicha membrana1, se obtendría una respuesta de tipo pasabanda.
Este comportamiento de la membrana basilar puede modelarse, con un grado de aproximación razonable, como una línea de transmisión no uniforme, representada en la Fig. III.11.
Cada etapa en paralelo representa un segmento corto de la membrana basilar. La corriente suministrada por la fuente corresponde a la velocidad del estribo. Los inductores en serie y en paralelo representan las masas del fluido y de segmentos de la membrana basilar, respectivamente; los condensadores representan la rigidez de la membrana, y se asume que su valor varía exponencialmente según la posición. Las resistencias representan pérdidas en la membrana.

Fig. III.11. Representación de la membrana basilar como una línea de transmisión.
Este modelo pasivo presenta varios inconvenientes: no considera fenómenos activos y no lineales de la membrana, no es capaz de generar una respuesta pasabanda tan estrecha como las observadas experimentalmente en tejidos vivos y, además, no toma en cuenta el hecho de que la membrana basilar es una estructura en tres dimensiones [5]. A pesar de ello, permite representar fácilmente los fenómenos de resonancia y de ondas viajeras.
En capítulos posteriores se discute un modelo análogo al anterior pero que resulta más útil en la elaboración de modelos perceptuales, en el cual se representa el efecto de la membrana basilar como el de un banco de filtros pasabanda. Si bien los parámetros que definen dicho banco de filtros se obtendrán a partir de consideraciones psicoacústicas, y no físicas o fisiológicas, se debe tener en mente que tal modelo está basado en las propiedades físicas observables de la membrana basilar y del oído interno en general.
III.6.Mecanismo de transducción
III.6.1.Interacción entre las membranas basilar y tectorial
El proceso de transducción o conversión de señal mecánica a electroquímica se desarrolla en el órgano de Corti, situado sobre la membrana basilar.
Las vibraciones de la membrana basilar hacen que ésta se mueva en sentido vertical. A su vez la membrana tectorial, ubicada sobre las células ciliares (los transductores), vibra igualmente; sin embargo, dado que los ejes de movimiento de ambas membranas son distintos, el efecto final es el de un desplazamiento "lateral" de la membrana tectorial con respecto a la membrana basilar.
Como resultado, los cilios de las células ciliares externas se "doblan" hacia un lado u otro (hacia la derecha, en la Fig. III.12, cuando la membrana basilar "sube").
En el caso de las células internas, aun cuando sus cilios no están en contacto directo con la membrana tectorial, los desplazamientos del líquido y su alta viscosidad (relativa a las dimensiones de los cilios) hacen que dichos cilios se doblen también en la misma dirección [9].

Fig. III.12. Desplazamiento relativo de las membranas basilar y tectorial.
III.6.2.Células ciliares y potenciales eléctricos
La diferencia fundamental entre los dos fluidos de la cóclea, la perilinfa y la endolinfa, estriba en las distintas concentraciones de iones en los dos fluidos. De esta manera, la endolinfa se encuentra a un potencial eléctrico ligeramente positivo (ver Fig. III.13) respecto a la perilinfa [3].

Fig. III.13. Potenciales eléctricos en el órgano de Corti y los fluidos de la cóclea.
Por otro lado, los movimientos de los cilios en una dirección determinada (hacia la derecha, en la Fig. III.12) hacen que la conductividad de la membrana de las células ciliares aumente [8]. Debido a las diferencias de potencial existentes, los cambios en la membrana modulan una corriente eléctrica que fluye a través de las células ciliares.
La consiguiente disminución en el potencial interno de las células internas provoca la activación de los terminales nerviosos aferentes, generándose un impulso nervioso que viaja hacia el cerebro. Por el contrario, cuando los cilios se doblan en la dirección opuesta, la conductividad de la membrana disminuye y se inhibe la generación de dichos impulsos.
Se pueden destacar dos aspectos de este proceso de transducción: primero, que la generación de impulsos nerviosos es un fenómeno probabilístico; segundo, que el proceso se comporta como un rectificador de media onda [2], puesto que la probabilidad de activación de las fibras nerviosas "sigue" a las porciones "positivas" de la señal sonora (equivalentes a desplazamientos hacia "arriba" de la membrana basilar, en la Fig. III.12), mientras que se hace cero en las porciones "negativas" de la onda.
III.6.3.Interacción entre células ciliares internas y externas
Como se dijo en la sección III.4.1, las fibras aferentes están conectadas mayormente con las células ciliares internas, por lo que es posible concluir con certeza que éstas son los verdaderos "sensores" del oído. Por el contrario, el papel de las células ciliares externas (más numerosas que las internas) era objeto de especulaciones hasta hace pocos años [8].
Recientemente se ha comprobado que dichas células no operan como receptores, sino como "músculos" [9], es decir, como elementos móviles que pueden modificar las oscilaciones en la membrana basilar.
La actuación de las células ciliares externas parece ser la siguiente [8] [9] [3]: para niveles de señal elevados, el movimiento del fluido que rodea los cilios de las células internas es suficiente para doblarlos, y las células externas se saturan. Sin embargo, cuando los niveles de señal son bajos, los desplazamientos de los cilios de las células internas son muy pequeños para activarlas; en este caso, las células externas se "alargan", aumentando la magnitud de la oscilación hasta que se saturan.
Este es un proceso no lineal de realimentación positiva de la energía mecánica, de modo que las células ciliares externas actúan como un control automático de ganancia, aumentando la sensibilidad del oído.
Este nuevo modelo del mecanismo de transducción nos indica que el conjunto formado por la membrana basilar y sus estructuras anexas forman un sistema activo, no lineal y con realimentación [3], y permite explicar dos fenómenos asociados al oído interno: el "tono de combinación", generado a partir de dos tonos de distinta frecuencia por un elemento no lineal que contiene un término cúbico [4], y las "emisiones otoacústicas", las cuales consisten en tonos generados en el oído interno en forma espontánea o estimulada [10], y que pueden llegar a ser audibles.
III.6.4.Selectividad en frecuencia de la cóclea
Debido a la acción de filtraje de la membrana basilar, cada célula transductora procesa una versión del estímulo sonoro filtrada de modo diferente [9]. Esta acción de filtraje de la membrana basilar por sí sola equivale a la de filtros cuya respuesta en frecuencia es relativamente "ancha". Ahora bien, la realimentación positiva provocada por las células ciliares externas contribuye a aumentar la selectividad del sistema auditivo.
Esto puede comprobarse midiendo la respuesta de una única fibra nerviosa ante variaciones en la frecuencia y la amplitud del estímulo sonoro [8]; las curvas de sintonía así obtenidas indican una respuesta de tipo pasabanda mucho más angosta que la debida al efecto de la membrana basilar como elemento pasivo.
Adicionalmente, experimentos recientes han permitido determinar que la selectividad del oído interno es virtualmente idéntica a la selectividad del sistema auditivo en su totalidad, estimada por métodos psicoacústicos [8].
Equilibrio
Los canales semicirculares y el vestíbulo están relacionados con el sentido del equilibrio. En estos canales hay pelos similares a los del órgano de Corti, y detectan los cambios de posición de la cabeza.
Los tres canales semicirculares se extienden desde el vestíbulo formando ángulos más o menos rectos entre sí, lo cual permite que los órganos sensoriales registren los movimientos que la cabeza realiza en cada uno de los tres planos del espacio: arriba y abajo, hacia adelante y hacia atrás, y hacia la izquierda o hacia la derecha. Sobre las células pilosas del vestíbulo se encuentran unos cristales de carbonato de calcio, conocidos en lenguaje técnico como otolitos y en lenguaje coloquial como arenilla del oído. Cuando la cabeza está inclinada, los otolitos cambian de posición y los pelos que se encuentran debajo responden al cambio de presión. Los ojos y ciertas células sensoriales de la piel y de tejidos internos, también ayudan a mantener el equilibrio; pero cuando el laberinto del oído está dañado, o destruido, se producen problemas de equilibrio. Es posible que quien padezca una enfermedad o un problema en el oído interno no pueda mantenerse de pie con los ojos cerrados sin tambalearse o sin caerse.
Descargar
| Enviado por: | Lion Grind |
| Idioma: | castellano |
| País: | España |
Todos los derechos reservados.