Biología, Botánica, Genética y Zoología
Biocatalizadores
taBLA DE CONTENIDOS
RESUMEN
Spanish
Los enzimas son catalizadores muy potentes y eficaces, químicamente proteínas, actúan en pequeña cantidad y se recuperan indefinidamente. No realizan reacciones que sean energéticamente desfavorables, no modifican el sentido de los equilibrios químicos, sino que aceleran su consecución. El sustrato se une a la enzima a través de numerosas interacciones débiles como son: puentes de hidrógeno, fuerzas electrostáticas, hidrófobas, etc, en un lugar específico, el centro activo.
Algunas enzimas actúan con la ayuda de estructuras no proteícas. En función de su naturaleza se denominan:
1.Cofactor: Cuando se trata de iones o moléculas inorgánicas.
2.Coenzima: Cuando es una molécula orgánica.
Las coenzimas pueden participar también covalentemente en las reacciones. La conformación de las proteínas depende, en parte, de sus cargas eléctricas, habrá un pH en el cual la conformación será la más adecuada para la actividad catalítica, pH óptimo. La temperatura òptima donde la actividad catalítica es máxima; por encima de esta temperatura, el aumento de velocidad de la reacción debido es contrarrestado por la pérdida de actividad catalítica debida a la desnaturalización térmica, y la actividad enzimática decrece rápidamente hasta anularse.
Exiten mecanismos para lo cual los procesos metabólicos son regulables via enzimas; el intento es el de caracterizar la totalidad de modelos de regulación de la actividad enzimática. Las enzimas resultan reguladas por la modificación de su cantidad o de su calidad. La modificación de la cantidad de la enzima se realiza por mecanismos genéticos, tales como:
La inducción enzimática
La represión enzimática
Al ocupar el sustrato el sitio activo, se produce una transformación de la enzima; ocurre el ajuste inducido, se produce la catálisis enzimática. La modificación de la calidad de una enzima significa cambios de su actividad catalítica. Las modificaciones de la actividad enzimática descansan en 5 mecanismos fundamentales:
Cascada enzimática.
Modificación Covalente.
Modificación Alostérica.
Isoenzimas.
Zimógenos
PALABRAS CLAVES : ENZIMOLOGÌA, SUSTRATO, MECANISMOS DE REACCION, ACTIVIDAD ENZIMÀTICA, ALOSTERISMO
English
The enzimas they are catalizadores very powerful and efficient, chemically proteins, they act in small quantity and they recuperate indefinitely. Not they carry out reactions that be energéticamente unfavorable, not they modify the sense of the chemical equilibria, but they accelerate their attainment. The sustrato joins with the enzima through numerous weak interactions as they are: bridges of hydrogen, strengths electrostáticas, hidrófobas, etc, in a specific place, the active center.
Some enzimas they act with the aid of structures not proteícas. In function of their nature they are called:
1.Cofactor: When is a matter of iones or moléculas inorganic.
2.Coenzima: When is a molécula organic.
The coenzimas they can participate also covalentemente in the reactions. The conformación of the proteínas depends, in part, of its electric loads, there will be a pH in which the conformación it will be more more adequate for the activity catalítica, pH optimum. The temperature òptima where the activity catalítica is maximum; over this temperature, the increase of velocity of the proper reaction is contrarrestado by the loss of activity catalítica due to the thermal denaturalization, and the activity enzimática decreases quickly to being annulled.
There is mechanisms for which the processes metabólicos are adjustable via enzimas; the intent is that of characterizing the regulation models totality of the activity enzimática. The enzimas they turn out to be regular by the modification of their quantity or of their quality.
The modification of the quantity of the enzima is carried out for mechanisms genéticos, such as:
The inducción enzimática
The represión enzimática
Al to occupy the sustrato the active place, a transformation is produced of the enzima; occurs the adjustment induced, is produced the catálisis enzimática. The modification of the quality of an enzima signifies changes of its activity catalítica. The modifications of the activity enzimática rest in 5 fundamental mechanisms: Waterfall enzimática. Modification Covalente. Modification Alostérica. Isoenzimas.Zimógenos
Key Words : ENZIMOLOGÌA, SUSTRATO, MECHANISMS OF REACCION, ACTIVITY ENZIMÀTICA, ALOSTERISMO
CARACTERÍSTICAS ESENCIALES DE LOS BIOCATALIZADORES
Biocatalizador quiere decir “catalizadores de la vida”, los biocatalizadores actúan sobre los seres vivos. Aquí entra el concepto de las enzimas. Las enzimas son proteínas que catalizan reacciones químicas en los seres vivos, son biocatalizadores porque están dentro de las células y producen reacciones biológicas. Los enzimas están en el metabolismo de los seres vivos, es decir, sustancias que, sin consumirse en una reacción, aumentan notablemente su velocidad. No hacen factibles las reacciones imposibles, sino que solamente aceleran las que espontáneamente podrían producirse. Ello hace posible que en condiciones fisiológicas tengan lugar reacciones que sin catalizador requerirían condiciones extremas de presión, temperatura o pH. Las enzimas usualmente trabajan a 37°C y pH=7 aunque hay que recalcar que hay enzimas que trabajan a condiciones extremas dentro del mismo organismo, sobre todo condiciones de pH por ejemplo las que actúan en el estomago que trabajan a pH´s menores a 2.5.
La velocidad de reacción al utilizar enzimas disminuye debido a que se necesita menos energía de activación para que se lleve a cabo la reacción, esto se entiende de mejor manera en el siguiente gráfico:
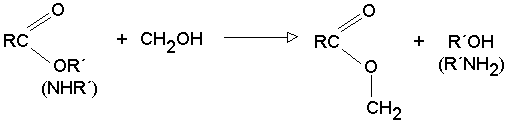
fig. 1 Energia vs. Sentido de la reaccion
Se ha dado el caso de que al aislar una enzima que en el cuerpo trabaja a pH=7 y hacerla reaccionar a otro valor de pH en laboratorio, esta tiene un mejor desempeño, es decir que existen enzimas en el organismo que no trabajan a el 100% de sus capacidades, por esa razón se afirma que nosotros somos seres que aun nos falta evolucionar para aprovechar la totalidad de la capacidad de una enzima.
El nombre de enzima, que fue propuesto en 1867 por el fisiólogo alemán Wilhelm Kühne (1837-1900), deriva de la frase griega enzymð, “en” significa “dentro” y “zyme” quiere decir “levadura”.
Como ya dijimos, las enzimas tienen una acción muy especifica y se clasifican en varias categorías: hidrolíticas, oxidantes, isómeras, de transferencia, reductoras, etc. dependiendo del tipo de reacción que controlen. Las enzimas hidrolíticas aceleran las reacciones en las que una sustancia se rompe en componentes más simples por reacción con moléculas de agua. Las enzimas oxidativas, conocidas como oxidasas, aceleran las reacciones de oxidación, y las reductoras las reacciones de reducción en las que se libera oxígeno. Las enzimas de transferencia actúan cuando se necesita la transferencia de grupos funcionales, cambian o sustituyen un grupo por otro. Las enzimas conocidas como ligasas son imprescindibles en la formación de enlaces con ruptura de ATP.
Cada enzima actúa sobre un sustrato, se muestra a manera de ejemplo esta tabla:
Tabla 1 Relacion Enzima-Sustrato
| Enzima | Sustrato |
| Sacarasa | Sacarosa |
| Lipasa | Lípidos |
| Proteasa | Proteínas |
| Carbohidrasa | Carbohidrato |
El sustrato se une a una región concreta del enzima, llamada centro activo. El centro activo comprende un sitio de unión formado por los aminoácidos que están en contacto directo con el sustrato y un sitio catalítico, formado por los aminoácidos directamente implicados en el mecanismo de la reacción. Las fuerzas que actúan en las uniones son los puentes hidrógeno, las interacciones electrostáticas y las fuerzas de VanDerWalls.
Una vez formados los productos el enzima puede comenzar un nuevo ciclo de reacción
El siguiente esquema muestra de que estamos hablando:
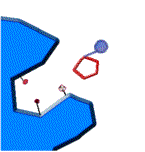
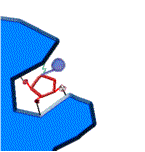
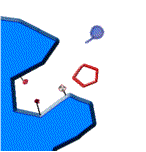
1.-la enzima y su sustrato 2.- unión al centro activo 3.-formación de productos
fig. 2 Proceso de reacción enzimática
Las enzimas, a diferencia de los catalizadores inorgánicos catalizan reacciones específicas. Sin embargo hay distintos grados de especificidad.
La enzima sacarasa por ejemplo, es muy específica, rompe el enlace ß-glucosídico de la sacarosa o de compuestos muy similares o análogos. Así, para el enzima sacarasa, la sacarosa es su sustrato natural, mientras que la maltosa y la isomaltosa son sustratos análogos. La enzima actúa con máxima eficacia sobre el sustrato natural y con menor eficacia sobre los sustratos análogos. Entre las enzimas poco específicas están las proteasas digestivas como la quimotripsina, que rompe los enlaces amida de proteínas y péptidos de muy diversos tipos.
Las enzimas logran que la velocidad de reacción se aumente de 100 a 1000 veces, se dice que son inteligentes porque saben cuando comenzar y cuando terminar. Hay enzimas que liberan energía para la contracción cardiaca y la expansión y contracción de los pulmones. Muchas facilitan la conversión de azúcar y alimentos en distintas sustancias que el organismo precisa para la construcción de tejidos, la reposición de células sanguíneas y la liberación de energía química para mover los músculos.
Existen enzimas como la tripsina y la pepsina que poseen una propiedad muy peculiar denominada autocatálisis que les permite originar su propia formación a partir de un precursor inerte denominado zimógeno. Como consecuencia, estas enzimas se pueden reproducir en un tubo de ensayo.
Las enzimas son muy eficaces. Cantidades pequeñas de una enzima pueden realizar a bajas temperaturas lo que podría requerir reactivos violentos y altas temperaturas con métodos químicos ordinarios. Por ejemplo, unos 30 g de pepsina cristalina pura son capaces de digerir casi dos toneladas métricas de clara de huevo en pocas horas. Una enzima no modifica el sentido de los equilibrios químicos
Efecto de la temperatura en la actividad enzimática
Es de conocimiento general que el aumento de la temperatura puede acelerar una reacción, las enzimas son inestables cuando se calientan.
El punto óptimo representa el máximo de actividad. A temperaturas bajas, las enzimas se hallan "muy rígidas" y cuando se supera un valor considerable (mayor de 50:) la actividad cae bruscamente porque, como proteína, el enzima se desnaturaliza. El punto optimo se aprecia claramente en el gráfico.
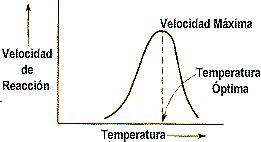
fig. 3 Efecto de la temperatura sobre la actividad enzimatica
Efecto del pH en la actividad enzimática
Los enzimas poseen grupos químicos ionizables (carboxilos -COOH; amino -NH2; tiol -SH; imidazol, etc.) en las cadenas laterales de sus aminoácidos. Según el pH del medio, estos grupos pueden tener carga eléctrica positiva, negativa o neutra. Como la conformación de las proteínas depende, en parte, de sus cargas eléctricas, habrá un pH en el cual la conformación será la más adecuada para la actividad catalítica. Este es el llamado pH óptimo.
El centro activo puede contener aminoácidos con grupos ionizados que pueden variar con el pH. La ionización de aminoácidos que no están en el centro activo puede provocar modificaciones en la conformación de la enzima. El sustrato puede verse afectado por las variaciones del pH.
Algunas enzimas presentan variaciones peculiares. La pepsina del estómago, presenta un óptimo a pH=2, y la fosfatasa alcalina del intestino un pH= 12
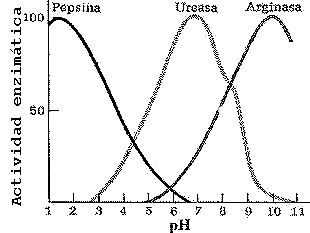
fig. 4 Efecto del pH sobre la actividad de la enzima
La mayoría de los enzimas son muy sensibles a los cambios de pH. Desviaciones de pocas décimas por encima o por debajo del pH óptimo pueden afectar drásticamente su actividad. Así, la pepsina gástrica tiene un pH óptimo de 2, la ureasa lo tiene a pH 7 y la arginasa lo tiene a pH 10 (Figura de la izquierda). Como ligeros cambios del pH pueden provocar la desnaturalización de la proteína, los seres vivos han desarrollado sistemas más o menos complejos para mantener estable el pH intracelular: se conocen como amortiguadores fisiológicos.
Efecto de los cofactores sobre la actividad enzimática
La actividad catalítica de una enzima está determinada sobre todo por su secuencia de aminoácidos y por la estructura terciaria, es decir, la estructura de plegamiento tridimensional de la macromolécula. Muchas enzimas precisan para su función la presencia de un ion o una molécula que recibe el nombre de cofactor.
Los cofactores son iones esenciales entre los cuales tenemos:
Iones activadores por ejemplo el Fe++ el cual puede entrar y salir de la enzima, o sea, no es parte estructural de la misma, solo se da una unión fuerte entre la enzima y el ion
Iones metálicos son los que en la enzima aun antes de que se estructure la misma, o sea, este ion metálico pasa a ser parte estructural de la enzima
Cabe señalar que los iones no solo puede ser Fe++, sino también Mn++, Mg++, Zn++, etc.
En el ejemplo del gráfico siguiente se utiliza como cofactor Co++ (Co++ en rojo y la enzima en verde).
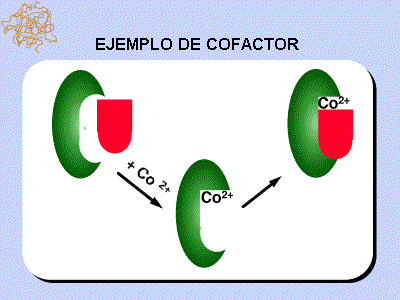
fig. 5 Cobalto como Cofactor
Casi un tercio de las enzimas conocidos requieren cofactores. Cuando el cofactor es una molécula orgánica se llama coenzima. Muchos de estas coenzimas se sintetizan a partir de vitaminas.
Cuando los cofactores y las coenzimas se encuentran unidos covalentemente a la enzima se llaman grupos prostéticos. La forma catalíticamente activa de la enzima, es decir, la enzima unida a su grupo prostético, se llama holoenzima. La parte proteica de una holoenzima (inactiva) se llama apoenzima, de forma que:
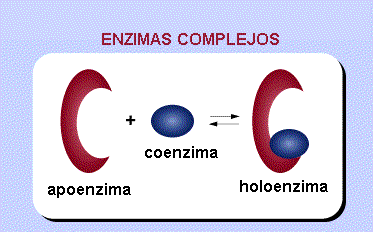
fig. 6 Holoenzima
En la figura inferior podemos observar una molécula de hemoglobina (proteína que transporta oxígeno) y su coenzima (el grupo hemo).
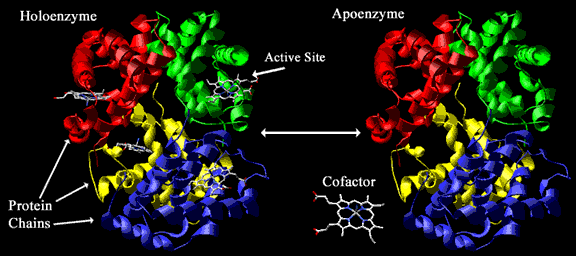
fig. 7 Apoenzimas, grupos prostéticos y Holoenzimas
En general los cofactores son sustancias no proteicas que se pegan a la proteína para que estas tengan actividad biológica.
Algunas VITAMINAS son necesarias para la actuación de determinadas enzimas, ya que funcionan como coenzimas que intervienen en distintas rutas metabólicas, por ello, la deficiencia en una vitamina puede originar importantes defectos metabólicos y realmente las deficiencias producidas por la falta de vitaminas responde más bien a que no se puede sintetizar una determinada enzima en la que la vitamina es la coenzima. En la tabla adjunta se puede apreciar algunas de las deficiencias causadas por este efecto:
Tabla2. Funciones de las vitaminas y enfermedades carenciales
| VITAMINAS | FUNCIONES | Enfermedades carenciales |
| C (ácido ascórbico) | Coenzima de algunas peptidasas. Interviene en la síntesis de colágeno | Escorbuto |
| B1 (tiamina) | Coenzima de las descarboxilasas y de las enzima que transfieren grupos aldehidos | Beriberi |
| B2 (riboflavina) | Constituyente de los coenzimas FAD y FMN | Dermatitis y lesiones en las mucosas |
| B3 (ácido pantotínico) | Constituyente de la CoA | Fatiga y trastornos del sueño |
| B5 (niacina) | Constituyente de las coenzimas NAD y NADP | Pelagra |
| B6 ( piridoxina) | Interviene en las reacciones de transferencia de grupos aminos. | Depresión, anemia |
| B12 (cobalamina) | Coenzima en la transferencia de grupos metilo. | Anemia perniciosa |
| Biotina | Coenzima de las enzimas que transfieren grupos carboxilo, en metabolismo de aminoácidos. | Fatiga, dermatitis... |
Como norma, las enzimas no atacan a las células vivas. Sin embargo, tan pronto muere una célula, ésta es digerida por enzimas que rompen sus proteínas. La resistencia de las células vivas se debe a la incapacidad de las enzimas de atravesar la membrana celular mientras las células tienen vida. Cuando la célula muere, su membrana se hace permeable y la enzima puede penetrar en la célula y destruir las proteínas en su interior. Algunas células contienen también enzimas inhibidoras, denominadas antienzimas, que evitan la acción de una enzima sobre un sustrato.
En este tema de las enzimas es importante hablar también de los inhibidores que son sustancias que estructuralmente se asemejan al sustrato de la enzima y que reacciona con la misma a velocidades muy bajas comparadas con la velocidad a la que reaccionaria el sustrato, o simplemente no reaccionan.
Cualquier sustancia que reduzca la velocidad de una reacción catalizada enzimáticamente se puede considerar que en un inhibidor. La inhibición de la actividad enzimática es uno de los mecanismos de regulación más importantes de las células vivas, y uno de los más importantes procedimientos de diagnóstico del enzimólogo. Los estudios de inhibición a menudo nos dicen algo cerca de la especificidad de un enzima, la estructura física y química del sitio activo y del mecanismo cinético de la reacción. En la vida cotidiana, los inhibidores enzimáticos pueden encontrarse enmascarados como drogas, antibióticos, preservativos, venenos y toxinas
Existen tres subdivisiones de los tipos más comunes de inhibición que pueden darse
Inhibición competitiva
Normalmente en el proceso de catalización enzimática de una reacción tenemos que se cumple este camino que viene a ser el transcurso usual de una reacción
E + S ![]()
ES ![]()
E + P.
Donde
E es la enzima
S es el sustrato
ES es el complejo enzima sustrato que es un punto intermedio de la reacción
P es el producto
Durante la inhibición competitiva entra en juego un inhibidor (I) que compite con el sustrato ya que se une a la enzima y baja su concentración efectiva
E + S + I -> EI + S + ES
Pero la cantidad de ES será muy poca comparada con la cantidad que hubiésemos obtenido de no haber intervenido un inhibidor. La cantidad del complejo ES puede incluso llegar a ser cero por lo que el uso del inhibidor puede resultar beneficioso en el caso de que lo que buscamos sea parar la reacción.
Inhibición incompetitiva
En este caso se presenta un inhibidor que actúa directamente en el complejo ES, o sea, se produce la reacción ES + I -> EIS no dándose el siguiente paso de la reacción que seria la obtención del producto deseado.
Inhibición no competitiva
Conocida también como inhibición mixta. En este caso el inhibidor actúa tanto sobre el complejo EIS como sobre la enzima pura, es decir que se producen estas dos reacciones simultáneamente:
E + S + I -> EI + S + ES
ES + I -> EIS
Cada reacción tendrá su propia constante de equilibrio y por tanto su propia velocidad de reacción. Si las dos reacciones tienen la misma afinidad nos encontramos ante el caso de inhibidores competitivos puros.
Uso industrial
A las enzimas se las puede utilizar en diferentes campos, por ejemplo:
En la industria alimenticia, textil, farmacéutica, análisis químicos, etc.
El problema de la inestabilidad de las enzimas y su alto costo, ha sido eliminado mediante el uso de enzimas inmovilizadas.
La fermentación alcohólica y otros procesos industriales importantes dependen de la acción de enzimas, sintetizadas por las levaduras y bacterias empleadas en el proceso de producción. Algunas enzimas se utilizan con fines médicos. En ocasiones son útiles en el tratamiento de zonas de inflamación local; la tripsina se emplea para eliminar sustancias extrañas y tejido muerto de las heridas y quemaduras.
Mediante técnicas de Biología Molecular se pueden obtener cepas mutantes que producirían la enzima en grandes cantidades, con lo que los costes del proceso se abaratarían.
Las enzimas se obtienen de microorganismos (bacterias, hongos o levaduras) seleccionados por screening, y posteriormente cultivados por fermentación (en matraz o reactor). A partir de los caldos de cultivo se procede a la purificación de la enzima que catalizaría la reacción de interés. Una vez purificada, se realizaría su caracterización, estudios cinéticos y de estabilidad en las condiciones de trabajo.
Por otra parte, se intentaría la clonación del gen que codifica para la enzima en Escherichia coli, lo que permitiría su máxima expresión y su obtención en grandes cantidades. Tanto la enzima salvaje como la enzima clonada, tras su purificación, se podría inmovilizar para su posterior recuperación y reutilización en el proceso. Las enzimas inmovilizadas podrían ser empleadas tanto en medio acuoso como en medio orgánico, según el tipo de biotransformación que se fuera a realizar.

fig. 8 Inmovilizacion de una enzima
La inmovilización es una tecnología que permite la obtención de derivados enzimáticos más estables que la enzima en solución, y que pueden ser reutilizados varias veces. También permiten el diseño de reactores de fácil manejo y control.
Utilización de tecnologías limpias en la síntesis de productos de química fina
Se ha desarrollado la tecnología necesaria para la síntesis selectiva, optimación y cambio de escala para la obtención de productos considerados de Química Fina.
Esta tecnología aborda el empleo de sistemas enzimáticos comerciales para sustituir a los catalizadores ácidos en la síntesis de ésteres, éteres, amidas, productos que se suelen utilizar en las industrias cosmética, farmacéutica, alimentación, aromas, etc.

fig. 9 Equipamiento utilizado en productos de química fina
Uno de los puntos claves de esta tecnología es la utilización de biocatalizadores comerciales, lipasas inmovilizadas. Comparado con los antiguos procesos, catalizados por ácidos minerales, resinas de intercambio, zeolitas o ácidos fuertes, esta tecnología permite tanto mejoras medio ambientales, como una disminución de los costes de operación y equipos de proceso.
Los productos desarrollados mediante esta tecnología han sido los siguientes:
Ésteres de alto peso molecular, para su aplicación en:
Alimentación: Aceite de Jojoba, Esperma de Ballena, DATEM, Monoglicéridos de oleico, esteárico.
Antioxidantes: Oleato de ascorbilo y Palmitato de ascorbilo.
Cosmética: Aceite de Jojoba, Monoglicéridos.
Farmacia: Cera Carnauba, Isopropil miristato
Ésteres de peso molecular medio, para su aplicación en:
Cosmética: Octanoato de n-octilo, miristato de miristilo, 2-etil-hexanoato de oleilo, Isopropil miristato.
Ésteres de bajo peso molecular, para su aplicación en:
Cosmética: Palmitato de isopropilo, oleato de isopropilo, oleato de metilo, oleato de etilo.
Surfactantes: 4-dihidroxibutanoato de oleilo.
Aminohidroxiésteres, para su aplicación en:
Farmacia: Intermedios de síntesis de fármacos.
Resolución de isómeros ópticos de mezclas racémicas, para su aplicación en el campo de la farmacología
ventaja
MECANISMOS DE ACCION CATALÍTICA
La catálisis enzimàtica y la especifidad depende de los grupos de superficie en el sitio activo de la enzima
Las enzimas aumentan la velocidad en la cual la reacciòn alcanza el equilibrio disminuyendo la energía de activaciòn, es decir, disminuyendo la energía libre de uno o màs intermediarios inestables de reacción. Las enzimas no alteran las posiciones de equilibrio de las reacciones que ellas catalizan. La energética de reacciones catalizadas y no catalizadas se compara esquemáticamente en la figura 6.1

Figura10. Energía libre de reacción vs. Coordenada de reacción
La especificidad y las propiedades catalìticas de una enzima son determinadas por los grupos químicos en una región de la superficie proteica, llamada sitio activo. El sitio activo de una proteína generalmente constituye menos del 5% de su àrea superficial. Las enzimas deben tener una conformación tal, que sus sitios activos como las cadena laterales responsables de la unión y de las reacciones de catálisis , pueden yuxtaponerse apropiadamente en tres dimensiones. El sitio activo siempre se encuentra en una grieta o hendidura y tiene dos funciones distintas : unión de sustrato (s) y catálisis . Generalmente, en cada función participan diferentes cadenas laterales de aminoácidos.
Las enzimas presentan una notable especificidad debido a sus encaje preciso entre sus sitios de unión y los sustratos. La unión del sustrato puede implicar enlaces iónicos, enlaces de H y enlaces de Van Der Waals. Los enlaces de van der waals contribuyen significativamente a la energía de unión sòlo cuando algunos estàn formados; es decir, cuando hay una considerable complementariedad molecular entre la enzima y el sustrato. Las enzimas se unen a los sustratos de tal modo que los àtomos participantes en el enlace que se va a formar o romper se orientan adecuadamente con respecto a los grupos catalìticos de la enzima. Las moléculas diferentes de sustrato que se unen al sitio activo de la enzima actúan como inhibidores competitivos de la catálisis.
Solamente unos pocos de los 20 aminoàcidos de las cadenas laterales participan directamente en la catálisis. En general, son cadenas laterales polares, cuyos participantes conocidos son Cis, His, Ser, Asp, Glu y Lis. Algunas veces tambièn estàn comprometidos los grupos amino y carboxilo terminales de la enzima. La mayorìa de las enzimas presentan una dependencia característica de la actividad por el pH, debido a la titulaciòn de uno o màs grupos cargados que participan en la catálisis (Fig. 6.2)

Figura11. Curva de activiada vs Ph de lisosima
Las cadenas laterales de aminoácidos no pueden catalizar todas las reacciones químicas requeridas por los sistemas biológicos. Por ejemplo, ninguna cadena lateral en buena aceptora de electrones. Por consiguiente, para la catálisis de algunas reacciones se requiere, además de las enzimas, moléculas pequeñas o iones metálicos (cofactores o coenzimas) con potencialidades químicas adicionales. Mientras que algunos cofactores se unen transitoriamente a la enzima durante la reacción, otros llamados grupos prostèticos se unen permanentemente a un aminoácido de la cadena lateral en el sitio activo de la enzima.
Catálisis covalente
Para entrar en explicaciones algo detalladas tenemos que tener presente que las enzimas tienen en sus grupos laterales cadenas en las que están presentes grupos carboxil, hidroxil, histadil y sulfhidril, estos son todos nucleofílicos. Las coenzimas pueden participar también covalentemente en las reacciones.

Tomemos el caso de la quimotripsina que es una enzima digestiva. La quimotripsina cataliza mediante hidrólisis una gran variedad de sustratos tanto esteres como amidas. También, mediante la utilización de sustratos que reaccionan lentamente o estudiando la razón a pH´s a los cuales la reacción es tortuosa, se ha acumulado bastante evidencia de la formación de un intermedio de asil serina.
Utilizando p-nitrofenil acetato, se puede seguir estequiométricamente el curso del p-nitrofenol. Si la formación del compuesto intermedio covalente es rápido, y la consecuente hidrólisis de la acetilenzima es lente, la tasa inicial de formación de p-nitrofenol debe ser mayor que la velocidad en estado estacionario después de que haya reaccionado una cantidad estequiométrica de un éster. Esta es la razón por la cual el fenol es mejor que el alcohol en estos casos. Los compuestos nucleofílicos competitivos pueden ser usados para proveer evidencia de un producto intermedio.

Cuando utilizamos amidas como sustrato, el segundo paso que es la hidrólisis de un éster, es mas rápida la formación del éster de la amida menos reactiva.
En soluciones de hidroxilamina, la velocidad de reacción de la amida es constante pero el producto varia con respecto a la concentración de hidroxilamina.

Se obtiene evidencia aun mas contundente mediante la cristalización de trimetil-acetil quimotripsina. Este voluminoso grupo acil es suficientemente apático en el segundo paso de la reacción como para ser aislado fácilmente.
Como vemos existen variedad de explicaciones incluso para un mismo experimento pero que representan una evidencia fehaciente de la existencia del sustrato intermedio que contiene a la enzima covalentemente enlazada.
Catálisis ácido-base
Comenzamos haciendo notar que el enlace carbón-oxígeno de los esteres es bastante fuerte. Para romper esteres en el laboratorio es necesario utilizar ácidos bastante fuertes o también bases igualmente fuertes. Los reactivos intermedios en estas reacciones no están disponibles en soluciones neutrales. Para una reacción enzimática, la transferencia de protones puede ser tan importante como crear y romper otros enlaces.

Como ya dijimos es posible hidrolizar un éster con reactivos a condiciones de pH realmente extremos, pero esto no es posible en una célula que permanece a pH moderado sin destruirla completamente.
A la catálisis mediante el ion hidrónio solo, sin la presencia de otros ácidos se le llama catálisis ácida especifica. En el caso de existir otros ácidos a parte de el ion hidrónio, la catálisis se denomina catálisis ácida general.
En estudios realizados con la amida acetilimidazol a pH´s entre 7 y 8, se encontró que la velocidad de reacción depende de la concentración de la solución buffer. El imidazol en el sitio activo de la enzima provee una catálisis básica general a las reacciones. El mecanismo de reacción de lo anotado seria el siguiente: el imidazol remueve el protón del hidrogeno enlazado con la molécula de agua de la solución. Como base remueve el protón del agua, el compuesto se convierte en nucleofílico, similar a un ion hidrogeno. La velocidad se incrementa. El ion imidazolio resultante entonces cataliza el rompimiento del enlace acil serina mediante el enlace del hidrogeno al oxigeno de la serina y transfiriendo un protón hacia el enlace ahora inestable del carbono con el oxigeno.
El agua es una buena base y también un buen ácido, por lo tanto es importante estudiar la catálisis enzimática en otros solventes, esto seria la catálisis ácido-base combinada.
La velocidad de hidrólisis del 2,3,4,6 tetrametilglucofuranosa en benceno fue estudiada con varios catalizadores. Se esperaba que esta reacción este sujeta a la catálisis ácido-base combinada. El fenol y la piridina catalizan la reacción, sin embargo, cuando el grupo básico acídico fue combinado en una molécula (2-pirodona), la catálisis fue mucho mas efectiva.

Comparado con la 2-pirodona el 2-aminofenol es un catalizador muy pobre. El correspondiente protón provoca un resultado desfavorable en la separación de las cargas.

Los efectos de los solventes no tienen gran influencia. Una amplia variedad de cadenas laterales de los aminoácidos hace posible que una enzima tenga un amplio rango de ambientes dentro de un mismo sitio activo.
Hemos visto varias interacciones de los sustratos con las enzimas en las reacciones, sin embargo, los cambios en el sustrato y en la interacción del sustrato con la enzima debe ocurrir como cambios de enlaces. Un gráfico estático de la catálisis de la enzima es insuficiente.
MECANISMOS DE REGULACIÓN DE LA ACTIVIDAD ENZIMÁTICA
Mecanismos para lo cual los procesos metabólicos son regulables via enzimas son ilustrados por selectos ejemplos. El intento es para caracterizar la totalidad de modelos de regulación de la actividad enzimática, esto implica que todos los necesarios reacciones enzima-catalizador procede en proporciones , respondiendo a los cambios en el medio ambiente externo como interno. Una célula u organismo puede ser definido como enfermo cuando responde incorrectamente o inadecuadamente a un interno o externo estímulo. El conocimiento de los factores que afectan una porción de las reacciones enzima-catalizador son esencialmente el conocimiento de los mecanismos de homeostasis en las celulas normales y el comprender los daños básicos moleculares.
Todas las reacciones químicas, incluidas enzima-catalizador , son para algunas reversibles extensiones, dentro de la vida de las células; sin embargo, reversiblemente no pueden ser obtenidas, porque los productos de las reacciones son prontamente removidas por adicion de reacciones enzima-catalizador. El flujo metabólico en la vida de las células en análogo a el flujo de agua en un tubo. A través del tubo se puede transferir agua en cualquier dirección, en la práctica el flujo es unidireccional. El flujo metabólico en la vida de las células es también unidireccional. El equilibrio verdadero, lejos de empezar las características de la vida, son aprovechadas únicamente cuando las células mueren. La vida de las células es un sistema dinámico de estado estable mantenido por un flujo unidireccional de metabolismos. (fig 11)

Figura 12. Una célula en satado estacionario.
En las células maduras, el significado de concentraciones del metabolismo ,permanece relativamente constante cerca en períodos de tiempo. La flexibilidad de los sistemas de estado estable son bien ilustrados en los cambios delicados y los balances por cada organismos mantenidos , la constancia de los cambios internos medio ambientales como comida, agua o una temperatura externa.
Mientras el conocimiento de los procesos regulares celulares en humanos es centrado a un entendimiento y terapia de enfermedades metabólicas, los eventos moleculares en regulación de muchos procesos metabólicos en mamíferos son pobremente entendidos. Esta claro que la regulación metabólica en diferentes mamíferos significa desde los superficiales fenómenos en bacterias. La regulación de los procesos metabólicos en bacterias serán discutidos porque proveen de una concepto “framework” para considerar las regulaciones en humanos.
El flujo neto de carbon atraves de alguna reacción enzima-catalizador puede ser influenciada por:
Por cambios de la cantidad absoluta de enzima.
Por alterar el lado de los reactantes de otra enzima
Por alterar la eficiencia catalítica de las enzimas
Las enzimas resultan reguladas por la modificación de su cantidad o de su calidad. La modificación de la cantidad de la enzima se realiza por mecanismos genéticos, tales como:
La inducción enzimática
La represión enzimática
A menudo puede estimularse la velocidad de síntesis de una enzima por exposición ( en la célula) a un inductor, un compuesto que es el sustrato para la enzima. Abundan ejemplos de inducción de enzimas, tanto en microorganismos como en animales superiores.
En la siguiente secuencia de gráficos, se explicara el proceso de ajuste inducido o inducción enzimática:
Se puede considerar la inhibición por retroalimentación como un mecanismo para reprimir la actividad de la enzima. El término represión enzimática es, sin embargo , el que se usa más frecuentemente para describir el efecto de un compuesto, llamado represor , que es el producto final de una serie de reacciones y que inhibe la síntesis de la enzima. Este mecanismo parece ser más eficiente para la regulación metabólica, pues permite a la célula utilizar sus aminoácidos para la síntesis de otras enzimas activas que necesita.

Figura 13.Regulación de la síntesis de proteìnas
La distinción entre una inzima inducida y la misma enzima cuando su presencia es normal en la célula ( una enzima constitutiva), probablemente es sólo de naturaleza cuantitativa, pues ambas parecen ser idénticas en todas los aspectos y la mayoría de las enzimas inducibles se encuentran en indicios antes de la inducción. Los efectos de la inducción no son permanentes, pues la eliminación del inductor conduce, después de varias generaciones de células, a una disminución en la cantidad de la enzima. Pueden actuar como inductores compuestos estrechamente afines en su estructura al substrato y la adición de un substrato puede conducir a la inducción de una serie de enzimas que intervienen en el metabolismo del sustrato -inducción en serie- pues los intermedios actuan también como inductores. El mecanismo genético de una célula de una célula tiene un gen apropiado para la formación de una enzima particular que puede ser inducida y el número de enzimas que pueden ser inducidas es limitado. En la siguiente figura se iluestra la inducción de galactosidasa en E. Coli, por diferentes inductores.

Figura 14 Aumento del N bacteriano.
Como una simple metación de gen puede cambiar una enzima inducidad en una enzima constitutiva , el mecanismo de formación de unas y otras debe ser el mismo. Aunque se ha sugerido que en una célula normal el papel del inductor lo desempeñe productos del metabolismo.
La modificación de la calidad de una enzima significa cambios de su actividad catalítica. Las modificaciones de la actividad enzimática descansan en 5 mecanismos fundamentales:
Cascada enzimática.
Modificación Covalente.
Modificación Alostérica.
Isoenzimas.
Zimógenos
Enzimas Alostéricas
El nombre de enzima alostérica fue dado por Jacques Monod, Jean Pierre Changeux y Jeffries Giman , del Instituto Pasteur de París, los cuales fueron los primeros en desarrollar una amplia teoría para la función de este tipo de enzimas reguladoras. El término alostérico significa - otro espacio - u - otra estructura - ; las enzimas alostéricas poseen, además del centro catalítico, el “ otro espacio ” al que se enlaza de modo reversible y no covalente el efector o modulador. En general, el centro alostérico es tan específico para la unión del modulador, como el centro catalítico lo es para la unión del sustrato.
El modelo de Michaelis - Mendel a tenido un gran efecto en el desarrollo de la química enzimática, sus virtudes son simples y de amplia aplicabilidad. Sin embargo, las propiedades cinéticas de muchas enzimas no pueden ser descritas por el modelo de Michaelis - Mendel. Un importante grupo consiste de las enzimas alostéricas, las cuales a veces presentan curvas sigmoidales de la reacción de velocidad, V, versus la concentración de sustrato, [S], en vez de la curva hiperbólica predicha por la ecuación de Michaelis - Mendel.
La primera enzima de la secuencia, la L-treonin-deshidratasa. es fuertemente inhibida por la L-isoleucina, que es el producto final,pero no lo es por ningún otro producto intermedio de la secuencia. Las características cinéticas de la inhibición por la isoleucina son atípicas; la inhibición no es ni competitiva con el sustrato, la L-treonina. ni tampoco no competitiva o acompetitiva. La isoleucina es muy específica como inhibidor, sin embargo otros aminoácidos o compuestos relacionados no inhiben. Este tipo de inhibición se designa de diversas maneras: inhibición por el producto final, inhibición fe^d-back y retroinhibición. El primer enzima de esta secuencia el cual es inhibido por el producto final, se llama enzima alostérico
Las enzimas alostérícos poseen,además del centro catalítico, el «otro espacio» al que se enlaza de modo reversible y no covalente el efector o modulador. En general, el centro alostérico es tan específico para la unión del modulador, como el centro catalítico lo es para la unión del sustrato. Algunos moduladores, como por ejemplo la L-isoleucina para la treonín-deshidratasa (fig. 9-18) son inhibidores, y por ello se les denomina moduladores inhibidores o negativos. Otros enzimas alostéricos pueden tener moduladores positivos o estimuladores. Cuando un enzima alostérico posee solamente un modulador específico. Ufe dice que es monovalente. Algunos enzimas alostéricos responden a dos o más moduladores específicos, cada uno de silos unido a un centro especifico del enzima; se dice entonces que son polivalentes
Las enzimas alostéricas poseen, pesos moleculares mucho mayores, son más complejos y con frecuencia más difíciles de purificar que los enzimas ordinarios, debido a que casi todos los enzimas alostéricos conocidos son oligómeros y poseen, por tanto, dos o más subunidades constituidas por cadenas polipeptidicas. por lo general en número par; hay algunos que contienen muchas cadenas. Los enzimas alostéricos muestran cierto número de propiedades anómalas. Algunos son inestables a O °C. pero estables a temperatura ambiente o a la del cuerpo.
Los enzimas alostéricos muestran dos tipos de control diferentes: heterotrópico y homotrópico, según la naturaleza de la molécula moduladora. Los enzimas heterotrópicos son estimulados o inhibidos por la molécula de un modulador o efector distinta de la de sus sustratos. Por otra parte, en los enzimas homotropicos el sustrato actúa también como modulador. Estos enzimas homotrópicos contienen dos o más centros de unión para el sustrato; la modulación de estos enzimas dependen de cuántos sean los centros del sustrato que estén ocupados. Sin embargo, una gran parte (si no todos) de los enzimas alostéricos son del tipo mixto homotrópico-heterotrópico sobre los cuales pueden actuar como moduladores, tanto el sustrato como algún otro metabolito.
En este tipo de enzimas, los ligamentos del sustrato a uno de los lados activos pueden afectar las propiedades del otro lado activo en la misma molécula de la enzima. Un posible resultado de esta interacción entre subunidades es que los ligamentos del substrato lleguen a ser cooperativos, lo que daría un curva sigmoidal de V versus [S]
La unión de una molécula de sustrato facilita la unión de la siguiente molécula de sustrato mediante un aumento en las afinidades de los sitios de unión vacantes. El fenómeno ha sido denominado “unión cooperativa” o “cooperatividad positiva” con respecto a la unión del sustrato o “respuesta homotrópica positiva”
Se ha propuesto dos modelos principales para las enzimas alostéricas. Son el modelo de “interacción secuencial ”y el modelo “ concentrado o de simetría”. Como su propio nombre indica, el model “secuencial ” supone unos cambios secuenciales o pregresivos en las afinidades de los sitios vacantes a medida que los sitios se van ocupando . El modelo “ concertado o de simetría” supone que la enzima preexiste como una mezcla en equilibrio de un oligómero de alta afinidad y un oligómero de baja afinidad. Los ligandos, incluido el sustrato, actúan desplazando el equilibrio a favor de uno u otro estado. Durante la transición, la conformación de todas las subunidades cambia al mismo tiempo.
MODELO SENCILLO DE INTERACCON SECUENCIAL
El model más sencillo de una enzima alostérica es una ampliación del modelo del “enzima flexible” o del “ajuste inducido” de Koshland. Este modelo supone que los cambios significativos en la conformación de una enzima pueden deberse a la unión de un sustrato u otro ligando . Estos cambios conformacionales pueden dar por resultado afinidades alteradas de los sitios vacantes . En efecto, cada molécula de sustrato que se une facilita la unión de la siguiente molécula de sustrato. La curva de velocidad resultante tiene una marcada fase de aceleración, seguida de la curvatura normal a medida que la enzima se aproxima a la saturación.
La ecuación de velocidad no se reduce a la ecuación de Henri-Michaelis-Menten.
Isoenzimas
En un contexto fundamental, el término isozima o isoenzima puede ser usado para abarcar la totalidad de ejemplos de distintas formas físicamente de la actividad catalítica. En la práctica, isozima un más restringido significado, es decir, los físicamente distintos y separadas formas de una enzima dada presente en diferentes tipos de células de una específica eucariota tanto como el comienzo humano. Isozimas estan comúnmente en el tejido de todos los vertebrados, insectos, plantas, y organismos unicelulares. Isoenzimas de numerosas hidrogenasas, oxidasas, transaminasas, fosfatasas, transfosforilasas, y enzimas proteolítias han sido reportadas.
En muchos sistemas multienzimáticos el producto final de la secuencia de reacciones puede actuar como un inhibidor especifico de un enzima situado al comienzo de la secuencia o muy próximo a él, lo cual determina que la velocidad de la secuencia completa de reacciones resulte condicionada por la concentración de producto final, en el estado estacionario. El ejemplo clásico es la secuencia multienzimática que cataliza la conversión de L-treonina en L-isoleucina, que procede en cinco etapas catalizadas enzimáticamente.
La existencia de estas formas múltiples puede detectarse y separarse mediante electroforesis en goles de los extractos celulares; puesto que están codificados por genes diferentes, los isozimas difieren en su composición aminoácida y por tanto en los valores de sus pH isoeléctricos.
El cuidadoso estudio cinético de los isozimas de la lactato deshidrogenasa ha demostrado que aunque todos ellos catalizan la misma reacción, difieren significativamente en sus valores de KM respecto a sus sustratos, particularmente respecto al piruvato, así como en los valores de Vmáx cuando el sustrato es el piruvato. El isozima M4 característico del músculo esquelético y de los tejidos embrionarios, posee un valor relativamente bajo de KM para el piruvato y una velocidad relativamente elevada en la reducción del piruvato a lactata El isozima H4, es característico del corazón y de otros músculos rojos, posee una KM relativamente elevada para el piruvato y lo reduce a una velocidad relativamente baja.
Hoy en día conocemos isozimas de un gran número de enzimas diferentes. Generalmente están constituidos por mezclas íntimamente asociadas de diferentes clases de cadenas polipeptídicas, en las que están combinadas-las propiedades cinéticas y de enlace específicas' aportadas por cada tipo de cadena. Muchos enzimas alostérícos se encuentran en forma de dos o más isozimas que varían en su sensibilidad respecto a sus moduladores alostéricos
En otros casos pueden encontrarse diferentes isozimas en los diversos compartimientos intracelulares. El estudio de los isozimas es de fundamental significación en la investigación de la base molecular de la diferenciación celular y de la morfogénesis. Muchas proteínas, y no sólo las que tienen actividad catalítica, pueden presentarse en formas múltiples en las células.
Figura 15. Reacción de lactato dehidrogenasa
Zimógenos
Estas proteinas son precursores de enzimas inactivas, relacionadas primariamente con la actividad proteolítica. Algunas proteinas son elaboradas desde sus células de origen en la forma de precursores inactivos de proteinas conocidos como “ proproteinas ”. Cuando las proteínas son enzimas,a las proproteinas se las conoce como “proenzimas ” o “zimógenos ”. La conversión de una proproteina a la proteina madura involucra un proceso conocido como limite, proteolisis selectiva. Para este proceso, la proproteina es convertida por uno o más proteolíticos clips sucesivos a una forma en la cual las actividades características de la proteina madura es expresada. Ejemplos de proteinas elaboradas como proproteinas son:
Pepsinógeno. Esta proteína, que se encuentra en la mucosa gástrica, tiene un peso molecular de 42 500 y se convierte por la propia pepsina en pepsina activa, con un peso molecular de 34 500. La conversión va acompañada de la separación de un quinto de la cadena de péptido . Uno de estos péptidos (peso molecular 3 100) es un inhibidor de la pepsina, pero se degrada subsiguientemente, por la enzima activa, a una mezcla de cuatro péptidos. Los otros cinco péptidos tienen pesos moleculares de 1 000 aproximadamente.
Tripsinógeno. Esta enzima es elaborada por el páncreas y se convierte en tripsina por una enzima en el jugo intestinal, o por la propia tripsina. La conversión comprende la separación de un solo hexapéptido que da tripsina activa, cuyo peso molecular es 23 800. Al igual que el pepsinógeno, el tripsinógeno es una sola cadena de polipéptido.
Los cambios que se producen en la estructura primaria como resultado de la escisión del péptido, se cree inducen alteraciones en la conformación de la molécula de proteína, de tal índole que dan lugar a la formación o a la exposición de los lugares catalíticamente activos, residuos de serina e histidina, que en la estructura primaria estaban muy alejados. Las tres histidinas están en la porción terminal amino de la cadena de polipéptido, mientras que la serina se encuentra en la región terminal carboxilo.
Procarboxipeptidasa. Esta enzima ha sido aislada del páncreas en dos formas: una molécula grande con un peso molecular de 87 000 y una más pequeña, con un peso molecular de 64000. La primera es procarboxipeptidasa A, con la cual se han efectuado muchos estudios. La activación de cualquiera de estos dos zimógenos, por tripsina, es causa de dos actividades:
A bajas concentraciones de tripsina se produce una enzima que, como la quimotripsina, actúa como una endopeptidasa, es sensible a la inactivación por diisopropilfluorofosfato e hidroliza enlaces adyacentes a residuos de tirosina o de fenilalanina, y
A concentraciones más altas de tripsina desaparece la actividad de endopeptidasa y aparece su actividad de exopeptidasa, por lo que hidroliza residuos de aminoácidos terminales C y esteres.
La activación de la enzima supone escisión de un enlace de péptido y liberación de carboxípeptidasa A activa, con un peso molecular de 34500. La enzima contiene un átomo gramo de cinc por peso molecular gramo y el metal es necesario para su actividad completa. El cinc está enlazado a un solo residuo de cisteína, al igual que a un grupo NH2 a de la asparagina terminal N. Cuando se separa el metal, se pierde la actividad enzimática, pero puede recuperarse por adición de Zn++ o por cualquiera de los metales del primer periodo de transición: Co++, Bi-M', Mn++ o Fe++. Cuando el Zn++ es reemplazado por los metales del grupo IIB, Cd++ y Hg4-4-, o por Pb++, se pierde la actividad de peptidasa, aunque se conserva la actividad de esterasa. En el lugar activo están involucrados dos residuos de tirosilo. El enlazamiento del substrato por la apoenzima exenta del metal, al igual que por la metaloenzima, y el requisito de la presencia del metal para la actividad enzimática, sugieren que existe diferencia entre los lugares de enlace y catalítico.
Quimotripsinógeno. Este zimógeno se encuentra en el páncreas y puede ser aislado en dos formas: A (o a) y B. La forma más común, quimotripsinógeno A, se convierte en varías formas activas de quimotripsina por un proceso que comprende eliminación sucesiva de fragmentos de péptido.
La quimotripsina tiene un peso molecular de 25000 y consta de tres cadenas de polípéptidos que están unidas entre sí por los enlaces de disulfurp. La activación de quimotripsinógeno A por tripsina supone una escisión del enlace de péptido entre arg-15 e leu-16.
BIBLIOGRAFÍA
-
BIOQUIMICA MEDICA; Dr. Abraham Mazur, Dr. Benjamín Harrow; Centro regional de ayuda técnica; méxico; Editorial interamericana S.A.; Primera edición; 1968 Pags.326-329
-
INTRODUCTION TO MODERN BIOCHEMISTRY; P.Karlson; Academic Press Inc., USA; Second printing; july 1963; Pags.72-75,140,146
-
AN INTRODUCCIÓN TO BIOCHEMICAL REACTION MECHANISMS; James N. Lowe, Lloyd L Ingraham; Prentice -Hall, Inc.; New-Yersey USA; 1974; Pags 10-21
-
HARPER´S REVIEW OF BIOCHEMISTRY, David W. Martin, Jr. , Peter A. Mayes, Victor W. Rodwell, and Daryl K. Granner; 20th Edition; Large Medical Publications, Los Altos Californis USA; 1983; Pags 59-61, 76-78, 88-94.
-
BIOQUÍMICA, William B. Wood, Impreso en Mèxico, 1977, Fondo educativo interamericano, S.A., Pags 125-139
Consultado el día Domingo 30 de diciembre de 2001
29
Al ocupar el sustrato el sitio activo, se produce una transformación de la enzima.
Ocurre el ajuste inducido, se produce
la catálisis enzimática.
La enzima recupera
su conformación inicial,el producto abandona el centro activo y la enzima queda libre.
Figura16. Detección de lactato en un electroferograma
Descargar
| Enviado por: | Beto |
| Idioma: | castellano |
| País: | Ecuador |
Todos los derechos reservados.